第四节 基因的转称与重组
遗传型变异还可通过两个不同性质细菌之间发生遗传物质的转移和重组而实现.在基因转移中,提供DNA的细菌为供体,而接受DNA的细菌是受体。基因转移后获得重组的子代,即具有供体与受体菌二者的主要特性。实现基因转移需要两个基本条件:一是全部或部分供体菌的基因相应进入受体菌;二是在受体菌中形成重组(杂交)的基因组。一般在亲缘关系相近,供、受体菌间容易发生重组,而无亲缘性的细菌间因基因组缺乏同源序列,不能或不易发生重组。重组子代菌产生的率很低,因此一般需要有选择条件使重组子代菌生长繁殖基因转移的方式和机理有几种不同形式。两个细菌细胞间可通过暂时的沟通(如接合);也可根本不接触,通过供体菌释放的DNA片段进入受体菌(如转化);也可通过噬菌体作媒介将供体菌的DNA片段包裹在其头部转移至受体菌(转导)。
一、转化
转化是受体菌直接摄取供体菌游离的DNA片段,通过与染色体重组,获得了供体菌的部分遗传特性。转化的DNA可以是细菌溶解后释放的,也可用人工方法抽提而获得。
转化首先在1928年由Griffith在肺炎球菌中发现,以后在葡萄球菌、嗜血杆菌也先后被发现。Griffith 的实验为:以无毒的Ⅱ型粗糙型(无荚膜)肺炎球菌注入小白鼠,并不引起动物死亡;以有毒力的Ⅲ型光滑型(有荚膜)肺
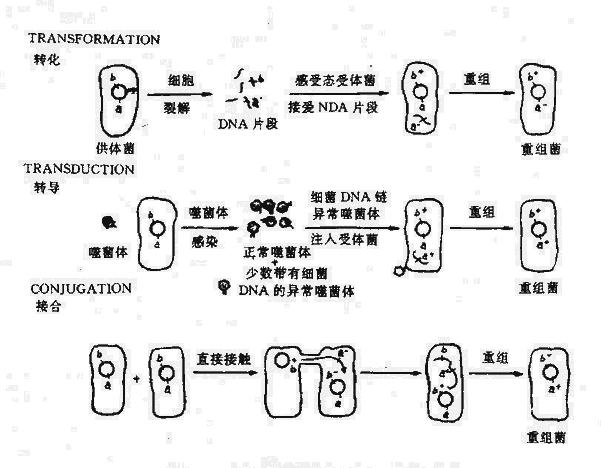
图5-4 细菌间的基因转移、转化、转导、接合
炎球菌注入小白鼠后,动物则死于全身性感染,以加热杀死的Ⅲ型光滑型肺炎双球菌注入小白鼠后,动物不死亡,亦分离不到Ⅲ型光滑型肺炎双球菌。如果将活的Ⅱ型粗糙型肺炎球菌与杀死的Ⅲ型光滑型肺炎球菌混合后注入小白鼠,结果动物发生全身性感染而死亡,自动物体内可分离到活的Ⅲ型光滑型肺炎球菌。以后直到1944年Avery才证实转化的物质是DNA,因可被DNA酶所破坏。以后的试管内培养条件下进行实验,发现细菌在摄取外源DNA时,需处于感受态(Competence)。肺炎球菌的感受态是在对数生长后期,约持续40分钟。进入受体菌的DNA片段需有与受体菌染色体上的同源核酸片段才能发生重组。当上述实验中Ⅲ型光滑型肺炎球菌产生荚膜的DNA片段与Ⅱ型粗糙型肺炎球菌的染色体DNA发生重组后,后者即可分裂产生具有Ⅲ型荚膜的光滑型有毒力的肺炎球菌子代。在自然条件下细菌通过转化获得外源性DNA发生遗传型变异的机会是存在的,但不一定多见。
二、转导
以噬菌体为媒介,把供细菌的基因转移到受体菌内,导致后者基因改变的过程称为转导。
当噬菌体在细菌中增殖并裂解细菌时,某些DNA噬菌体(称为普遍性转导噬菌体)可在罕见的情况下(约105~107次包装中发生一次),将细菌的DNA误作为噬菌体本身的DNA包入头部蛋白衣壳内。当裂解细菌后,释放出来的噬菌体通过感染易感细菌则可将供体菌的DNA携带进入受体菌内。如发生重组则受体菌获得了噬菌体媒介转移的供体菌DNA片段。这一过程称为普遍性转导。质粒也有可能被包入衣壳进行转导。不具有转移装置的质粒依赖噬菌体媒介进行转移,转导可转移比转化更大片段的DNA,转移DNA的效率较转化为高。
另一种转导称为局限性转导,指仅为特殊局限的一部分细菌DNA能被转导。只有温和噬菌体可进行局限性转导。当温和噬菌体进入溶原期时,则以前噬菌体形式整合于细菌染色体的一个部位。当其受激活或自发进入裂解期时,如果该噬菌体DNA在脱离细菌染色体时发生偏离,则仅为与前噬菌体邻近的细菌染色体DNA有可能被包装入噬菌体蛋白质衣壳内。因此局限性转导噬菌体所携带的细菌基因只限于插入部位附近的基因。由于局限性转导噬菌体常缺少噬菌体正常所需的基因,因此常需与野生型噬菌体共同感染细菌后的细菌中复制,这样才能将携带的基因转移至受体菌,并获得该段基因所决定的新特性的表达。
三、接合
两个通过直接接触,在暂时的沟通中进行基因转移的过程为接合。这一过程不是在所有细菌之间均可发生。只有那些具有F因子或类似F因子传递装置的细菌才能接合。接合中,有F因子的细菌相当于雌性菌。因此接合看作是细菌的有性生殖过程,又称为细菌杂交。
细菌的接合最早在大肠杆菌中发现,以后在其他菌中也观察到,主要见于革兰氏阴性菌。在电镜下可观察到细菌间借伸长的性菌毛进行接合。细菌能在接合中作为基因传递供体取决于致育因子(Fertility factor)又称F因子。这是最早发现的一种质粒。F因子编码在细菌表面产生性菌毛。F因子的特性为可以促进供体菌向受体菌传递色体DNA或质粒。F因子决定编码的性菌毛可在供体与受体菌间形成交通通连接结构,从而可使两个杂交细菌间形成胞浆内连接桥。F因子可以游离存大于胞浆内,也可与细菌染色体整合。如果F因子游离存在于胞浆内,接合时仅F因子DNA可通过胞浆的连接桥进入受体菌。然而F因子转移的特点为,从一个起始点开始,仅有一条DNA链进入受体菌,以后供体、受体菌分别以一条DNA链为模板,以滚环式复制另一条互补链,形成完整的双链F因子。这一特性使F因子与其他能通过接合传递的细菌质粒一样,在细菌群体中传播,类似引起传染,即原来的F+菌仍为F+,而F-受体菌可变成F-菌。
除F因子外,发现耐质粒R因子中有些亦可通过接合而传递,另一些则不能传递。R因子是1959年由日本学者所发现。他们对一批应用常用抗生素治疗无效的痢疾患者粪便中分离到的痢疾杆菌进行分析,发现细菌中有一种能同时耐几种抗生素的基因。这种基因存在于细胞浆中,可通过类似F因子的方式在细菌间传递。以后发现这类质粒中可通过接合转移者除有决定耐药性的r区段DNA外,还有传递区段(RTF,Resistance trarnsfer factor)。RTF决定性菌毛的形成,通过接合而传递。如果只有r区段而无RTF区段则不能过接合传递。必须经传递性质粒带动、噬菌体转导或以转化方式转入受体菌。
R因子决定细菌耐药性的问题是临床治疗中的大问题。R因子决定耐药性的机制,现已了解者为:1.质粒基因可编码产生各种纯化酶,如金黄色葡萄球菌耐药性质粒编码青霉素酶,耐氨苄青霉素的肠道杆菌质粒中编码能使β内酰胺环水解的酶。2.R因子通过控制一些细菌细胞膜的通透性,使四环素不能进入菌体。3.R因子通过阻止抗生素与细菌细胞内的作用部位(靶)结合,使细菌耐药。如红霉素通过与细菌核蛋白体结合而阻止蛋白质合成。R因子编码甲基酶,通过使核蛋白体上某些分子的甲基化,使结霉素不能与之结合而失去作用。由于R因子可通过接合的种、属不同的细菌间转移,因此有些痢疾杆菌即使未与药物接触过,但可自耐药的大肠杆菌获得R因子而耐药。目前有学者主张应及时了解医院内细菌的R因子质粒耐药图谱,轮流选用抗生素以达到较好的治疗效果。
除了上述各种基因转移的方式外,还发现了一类能在质粒之间或质粒与染色体之间自行转移位置的核苷酸序列,称为转座因子(Trnasposible elements)。其中最简单者仅有1,000个碱基对。只具有编码转移决定子的基因,称为插入顺序。还有一些分子量较大者为转座子。一般转座的DNA链末端有互补及倒置重复序列,从而一条单链即可自己形成环状结构。转座子插入细菌染色体后,因在插入部位影响了细菌染色体DNA的正常结构,可致细菌失去某些功能。如耐药基因。产生细菌霉素或某些酶的基因等。转座子携带的这些基因在即使与受体菌无核酸同源性的情况下仍可传递转移。因此转座子与质粒一样在构成致病性、耐药性菌中占有重要地位。