第三节 基因的复制与表达
生物的遗传物质基础是核酸(nucleic acid),它也是基因的基本结构,它们的化学组成分子结构符合遗传物质的稳定性、连续性及多样性的要求。
(一)核酸的化学组成
核酸结构的基本单位是核苷酸(nucleic acid),每个核苷酸由1个磷酸、1个五碳糖和1个碱基3部分组成。核酸有两类:一类是脱氧核糖核酸(DNA),DNA中的脱氧核糖核苷酸主要由4种碱基构成,即腺嘌呤(adenine,A)、鸟嘌呤(guanine,G)、胞嘧啶(cytosine,C)和胸腺嘧啶(thymine,T),此外,还有脱氧核糖(deoxyribose)和磷酸;另一类是核糖核酸(RNA),RNA分子中的核糖核苷酸主要由碱基A、G、C和尿嘧啶(uracil,U)构成,此外,还有核糖(ribose)和磷酸。
(二)DNA的分子结构
DNA分子是4种脱氧核苷酸经3’→5’磷酸二酯健聚合而成,所以了称为多核苷酸(polynucleotide)。DNA的一级结构是指4种核苷酸的连接及其排列顺序。1953年Watson和Cricd提出了DNA分子双螺旋结构模型,其要点是:DNA分子是由2条平行的多核苷酸链围绕同一中心轴构成的右手双螺旋结构(B型DNA)。多种芏酸的方向由核苷酸间的磷酸二酯健的走向决定,一条从5’→3’,另一条从3’→5’,两条链呈反向平行排列(antiparallel),彼此由氢键相连,G 与C配对(G≡C),A与T配对(A=T)。图3-3表明DNA的分子骨架。
图3-3 DNA双螺旋结构及碱基配对示意图
(A)部分SNA多核苷酸链,示邻近脱氧核苷酸由3’-5’磷酸二酯键连接;
(B)DNA互补的两条链;(C)DNA双螺旋模型
根据以上原则,只要确定了一条链中的碱基顺序,就可以相应在确定与它互补的另一条链上碱基的顺序,估计1个DNA分子大约有4千至40亿个核酸对,而各种碱基对排列顺序没有限制,即假定某一段DNA分子链有100个碱基对,则该段就有4100各不同的排列组合形式,即可有4100种不同性质的基因。现知,基因就是DNA分子链上的一个特定的区段,其平均大小约为1 000个碱基对。这说明对DNA分子贮存了大量正常和变异的遗传信息,满足了生物的遗传和多样性的要求,特别是通过DNA分子的准确复制,又可使遗传信息得到稳定和连续的传递。
(三)DNA的复制
基因的复制的以DNA复制为基础的。生物体的遗传特征表现为特定的核苷酸顺序,并以密码子的形式编码在DNA分子上。在细胞分裂过程中,通过DNA准确地自体复制(self-replication),把遗传信息从亲代传给子代,这样,DNA就能真正完成其作为遗传信息载体的使命,从而保证遗传物质的连续性和相对的稳定性。
由于DNA分子两条链的碱基是互补的,一条链上的核苷酸排列顺序可以由另一条链上的核苷酸排列顺序决定。DNA复制过程中,首先碱基间氢断裂,双螺旋解旋并松开,然后每条多核苷酸链各以自己为模板(template)吸收周围游离核苷酸,按碱基互补原则,进行氢键结合。在一些聚合酶作用下,合成新的互补的链,与原来模板单链并列盘旋在一起,形成了稳定的双螺旋结构(图3-4)。
这样新形成的2个DNA分子与原来 DNA分子的碱基顺序完全一样。每个子代DNA分子的一条链来自亲代DNA,另一条链则是新合成的,所以这种复制方式称为半保留复制(swmi-conservative replication).
图3-4 双螺旋DNA半保留复制过程P1,P2:DNA亲链;F1,F2:DNA子链;
示复制结果的两个DNA分子完全相同,每个新DNA分子均由亲链和子链组成
(四)基因的表达
所谓基因表达(gene expression)是指细胞在生命过程中,把储存在DNA顺序中遗传信息经过转录和翻译,转变成具有生物活性的蛋白质分子.生物体内的各种功能蛋白质和酶都是同相应的结构基因编码的(图3-5)。
1.转录过程 在RNA聚合酶的催化下,以DNA为模板合成mRNA的过程称为转录(transcription).在双链DNA中,作为转录模板的链称为模板链(template strand),或反义链(antisensestrand);而不作为转录模板的链称为编码链(coding strand),或有义链(sense strand).在双链DNA中与转录模板互补的一条DNA链即编码链,它与转录产物的差异仅在于DNA中T变为RNA中的U.在含许多基因的DNA双链中,每个基因的模板链并不总是在同一条链上,亦即一条链可作为某些基因的模板链的,也可是另外一些基因的编码链。
转录后要进行加工,转录后的加工包括:
(1)剪接:一个基因的外显子和内含子都转录在一条原始转录物RNA分子中,称为前mRNA(pre-mRNA),又称核内异质RNA(heterogenuous nuclear RNA,huRNA)。因此前mRNA分子既有外显子顺序又有内含子顺序,另外还包括编码区前面及后面非翻译顺序。这些内含子顺序必须除支而把外显子顺序连接起来,才能产生成熟的有功能的mRNA分子,这个过程称为RNA剪接(RNa splicing)。剪切发生在外显子的3’末端的GT和内含子3’ 末端与下一个外显子交界的AG处。
(2)加帽:几乎全部的真核 mRNa 端都具“帽子”结构。虽然真核生物的mRNA的转录以嘌呤核苷酸三磷酸(pppAG或pppG)领头,但在5’端的一个核苷酸总是7-甲基鸟核苷三磷酸(m7GpppAGpNp)。mNRA5’端的这种结构称为帽子(cap)。不同真核生物的mRNA具有不同的帽子。
mRNA的帽结构功能:①能被核糖体小亚基识别,促使mRNA和核糖体的结合;②m7Gppp结构能有效地封闭RNa 5’末端,以保护mRNA免疫5’核酸外切酶的降解,增强mRNA的稳定
(3)加尾:大多数真核生物的mRNA 3’末端都有由100~200个A组成的Poly(A)尾巴。Poly(A)尾不是由DNA编码的,而是转录后的前mRNA以ATP为前体,由RNA末端腺苷酸转移酶,即Ploy(A)聚合酶催化聚合到3’末端。加尾并非加在转录终止的3’末端,而是在转录产物的3’末端,由一个特异性酶识别切点上游方向13~20碱基的加尾识别信号AAUAAA以及切点下游的保守顺序GUGUGUG,把切点下游的一段切除,然后再由Poly(A)聚合酶催化,加上Poly(A)尾巴,如果这一识别信号发生突变,则切除作用和多聚腺苷酸化作用均显著降低(图3-5)。
图3-5 真核生物结构基因表达(DNA→RNA→蛋白质)流程图
mRNAPoly(A)尾的功能是:①可能有助mRNA从核到细胞质转运;②避免在细胞中受到核酶降解,增强mRNA的稳定性。
2.翻译过程真核细胞的转录以及加工都是细胞核内进行,但翻译过程则在细胞质中进行。
以mRNA作为模板,tRNA作为运载工具,在有关酶、辅助因子和能量的作用下将活化的氨基酸在核糖体(亦称核蛋白体)上装配为蛋白质多肽链的过程,称为翻译(translation),这一过程大致可分为3个阶段 (图3-6):
(1)肽链的起始:在许多起始因子的作用下,首先是核糖体的小亚基和mRNA上的起始密码子结合,然后甲酰甲硫氨酰tRNA(tRNA fMet)结合上去,构成起始复合物。通过tRNA的反密码子UAC,识别mRNA上的起始密码子AUG,并相互配对,随后核糖体大亚基结合到小亚基上去,形成稳定的复合体,从而完成了起始的作用。]
(2)肽链的延和长:核糖体上有两个结合点——P位和A位,可以同时结合两个氨酰tRNA。当核糖体沿着mRNA从5’→3’移动时,便依次读出密码子。首先是tRNAfMet结合在P位,随后第二个氨酰tRNA进入A位。此时,在肽基转移酶的催化下,P位和A位上的2个氨基酸之间形成肽键。第一个tRNA失去了所携带的氨基酸而从P位脱落,P位空载。A位上的氨酰tRNA在移
图3-6 翻译过程(蛋白质合成)图解
位酶和GTP的作用下,移到P位,A位则空载。核糖体沿mRNA 5’端向3’端移动一个密码子的距离。第三个氨酰tRNA进入A位,与P位上氨基酸再形成肽键,并接受P位上的肽链,P位上tRNA释放,A位上肽链又移到P位,如此反复进行,肽链不断延长,直到mRNA的终止密码出现时,没有一个氨酰tRNA可与它结合,于是肽链延长终止。
(3)肽链的终止:终止信号是mRNA上的终止密码子(UAA、UAG或UGA)。当核糖体沿着mRNA移动时,多肽链不断延长,到A位上出现终止信号后,就不再有任何氨酰tRNA接上去,多肽链的合成就进入终止阶段。在释放因子的作用下,肽酰tRNA的的酯键分开,于是完整的多肽链和核糖体的大亚基便释放出来,然后小亚基也脱离mRNA.
(4)翻译后加工(postranslational processing):从核糖体上释放出来的多肽需要进一步加工修饰才能形成具有生物活性的蛋白质。翻译后的肽链加工包括肽链切断,某些氨基酸的羟基化、磷酸化、乙酰化、糖基化等。真核生物在新生手肽链翻译后将甲硫氨酸裂解掉。有一类基因的翻译产物前体含有多种氨基酸顺序,可以切断为不同的蛋白质或肽,称为多蛋白质(polyprotein)。例如胰岛素(insulin)是先合成86个氨基酸的初级翻译产物,称为胰岛素原(proinsulin),胰岛素原包括A、B、C三段,经过加工,切去其中无活性的C肽段,并在A肽和B肽之间形成二硫键,这样才得到由51个氨基酸组成的有活性的胰岛素。
3.外显子与内含子表达过程中的相对性 从内含子与外显子的定义来看,两者是不能混淆的,但是真核生物的外显子也并非都“显”(编码氨基酸),除了tRNA基因和rRNA基因的外显子完全“不显”之外,几乎全部的结构基因的首尾两外显子都只有部分核苷酸顺序编码氨基酸,还有完全不编码基酸的外显子,如人类G6PD基因的第一外显子核苷酸顺序。
现在已发现一个基因的外显子可以是另一基因的内含子,所这亦然。以小鼠的淀粉酶基因为例,来源于肝的与来源于唾液腺的是同一基因。淀粉酶基因包括4个外显子,肝生成的淀粉酶不保留外显子1,而唾液腺中的淀粉酶则保留了外显子1的50bp顺序,但把外显子2与前后两段内含子一起剪切掉,经过这样剪接,外显子2就变成唾液淀粉酶基因中的内含子。
4.同一基因在不同组织能生成不同的基因产物 来源于不同组织的类似蛋白,可以由同一基因编码产生,这种现象首先是由于基因中的增强子等有组织特异性,它能与不同组织中的组织特异因子结合,故在不同组织中同一基因会产生不同的转录物与转录后加工作用。此外真核生物基因可有一个以一的poly(A)位点,因此能在不同的细胞中产生具有不同3’末端的前mRNA,从而会有不同的剪接方式。由于大多数真核生物基因的转录物是先加poly(A)尾巴,然后再行剪接,因此不同组织、细胞中会有不同的因子干预多聚腺苷酸化作用,最后影响剪接模式。降钙素(calcitonin)基因在不同组织中的表达可作为实例(图3-7)。
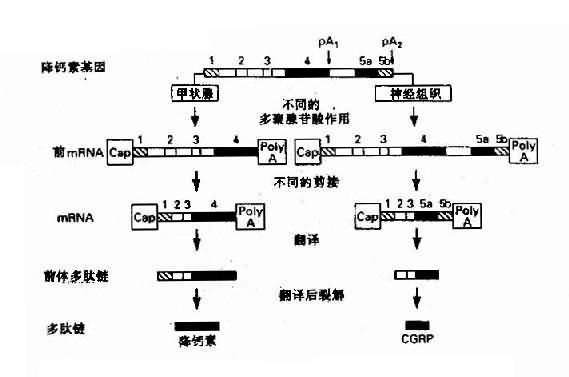
图3-7 降钙基因在不同的细胞中产生不同的激素
1:为非翻译顺序;2、3:为共同的编码外显子;
4:为降钙素外显子;5 a:为CGRP外显子;5b:为CGRP3’ 非翻译顺序;
pA1、pA2:为AATAAA加尾信号;CGRP:与钙素基因相关的多肽
降钙素基因(CALCAa多肽,定位于11p15.4)中甲状腺细胞中形成的前mRNA(短转录物),包含有非翻译顺序(1)、编码外显子(2)和(3)以及降钙素编码外显子(4)(包括部分非编码区),在转录物pA1位点即AAUAAA信号附近进行多聚腺苷酸化。而在下丘脑,其前mRNA(长转录物)中除了包含转录物的全部顺序外,还包含有与降钙素基因相关肽(calcitonin gene-related peptide, CGRP)的编码外显子(5a)和CGRP的3’末端非翻译顺序(5b),并在转录物pA2处进行poly(A)加工。可是,在长转录物的剪接过程中,外显子(3)的拼接点直接与CGRP编码外显子(5a)拼接点相连,从而删除降钙素的编码外显子(4),这样形成两种成熟的mRNA,分别翻译产生降钙素的前体和CGRP前体,然后通过酶促降产生降钙素CGRP这两种激素。