4��2 ̼ˮ�����������л
����4��2��1 ��������
����̼ˮ����Ҫ�������Dz����ա������Ĺ��̾���ˮ��Ĺ��̡���ѿ�ǡ����ǡ����ǡ���ѿ�;��Ƕ������������������Ķ��ǽ�����һ�֣���ԭ���Ƴ�ʳƷʱ�Ѳ������ˡ�
���������ӿ�ǻ��ʼ����ǻ������Һ����ø��ˮ�⽻���1��4�Ǹʼ���������ˮ���1��6���ռ������ڵĦ�1��6���ռ������������Ǻ�������ѿ�;��Ǻ���ѿ�ǡ�
����θ��û���������۵�ø����Һ����ø������PH��6.6��6.8����ʳ��û�б�θ���к���ǰ���ܳ�������һ��ʱ�䣬ʹ���ۺ͵;�����������һ���֡�
����С��������Һ�Ħ�-����ø�������ú���Һ����ø��ͬ����ֱ��������������ѿ�Ǻ���ѿ���ǣ�֧��������������ѿ�ǡ���ѿ���Ǽ���4��9��������������ɵĶ��Ц�1��6�ռ�����ѿ�;��ǡ�
������ճĤ��Ƥϸ����������ϸ����ÿһϸ��Լ��3000����ë����ë��Ŀռ����Ч�뾶Լ0.4nm��ֻ���������������ܹ�ͨ��������ëĤ�ϵ�ø��Ӧ��Ĥ�ϵ�ø�����֣��٦�1��4����ø���������Ƿ�������������һ��������������������ѿ��ø��ˮ����ѿ�;��ǵĦ�1��6���ռ���������ø���������ǣ��ܦ�-������ø���������ǡ�
���������������������У��ٳ�ǻ�ڵ�������������˫�Ǻ���ѿ�;��ǣ�����ëĤ�ϵ������������ǵ��ǡ�
������ë˫֬�ʲ�ģ��������øǶ��˫֬�ʲ��ڣ�����λ����Ĥ�⡣�������������ȫ��Ƕ��Ĥ�ڵ����䵥�ǵĵ����ʣ����������������ղ������̿��Ա����䵰������ϡ�
�������䵰���ڽ����������ǰ���Ƚ�ϳ�ǻ�ڵ�Na+���볦ǻ����ǻ��Na+������ϸ���ڣ��ͷŵ������С�Na+���볦ǻ����ǻ�е�Na+Ũ�ȱ�ϸ���ڸߣ��Ե�Ũ���ŵ���Ũ��Ҫ��������������������ATP��Ӧ���ǽ���ϸ����Լ��15%���س�ǻ��25%��ɢ��Ѫ��60%�뿿����Ĥһ�˵���Ĥ�ϵ���һ���嵰��϶��뿪ϸ������һ��ϲ���Na+���������������ǵ��ٶȱ������Ǵӳ�ǻ��������ϸ�����ٶȿ죬���������Dz���������ϸ����������Ӷ����������Ч�ʡ���ʳ�ӵ���ճ��²�ʱ��95%��̼ˮ���ﶼ�������ˡ�
���� 4��2��2�����
����С�����յ�̼ˮ������Ҫ�������ǡ����������������֣����ž����͵����ࡣ�����ǽ����ϸ���������ᷴӦ����������-6-���ᣬ����ϸ���ڵ�������Ũ�ȿ�ά���ڵ�ˮƽ��ʹ�����Dz��Ͻ����ϸ���������յ��������У�60%�����ڸ��ڴ�л���������ѭ�������ǺͰ������ڸ���ת��Ϊ�����ǡ�
�����������ڸ��ھ��ֽ��л�ṩ��������Ҫ������������ĺϳ���ԭ�����ڸ��ڣ����ж����ת���֬�����͵�֬����֯����������
�����ǵķֽ��л�������Σ�
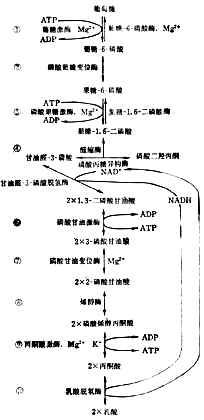
������1���ͽ������Ƿֽ�ĵ�һ�������ɱ�ͪ�ᡣ��һϵ�з�Ӧ��̼ˮ���������⾭��ĸ���������ɱ�ͪ����ͬ�����Գ�Ϊ�ͽ⡣����ͬ���Ƿ���ʱ��ͪ���һ�����������������ڣ�����ʱ��ͪ��������������̼��ˮ������ʱ��ԭ��������
�����ͽ��ʮ�����У�ͼ4-11����Ϊ��Ҫ�������ǵ�6��̼ԭ��һ��Ϊ������ʹ���������ữ�����ܶ��г������ӽ���ȵIJ��֣�Ȼ����ȥ����ɱ�ͪ�ᡣ
����������Ӧ����ϸ�����н��С�ÿһ�������ض���ø�μӡ��ڢ١�����������2����ATP��˵�������Ƿֽ��л��ʼʱҪ���������ƶ����ޡ�������������2����ATP�������ѵ���ͪ�ᣬ����2����ATP���ͽ�������������ࡣ
���� ϸ������NAD+��NADH�����������㶨�������е�NAD+�����NADH���ͽ�͵�ֹͣ��Ҫʹ���Ѽ�������������ͪ�ᣬ�����NADH����ת�Ƶ������������ȥ������ʱ�⾭������������ˮ������ʱ�⽻����ͪ��ʹ������⡣�������ʱ�뿪ϸ����Ѫ����������ȱ���߿ɷ�����л�����ж���
���� ��2����ͪ����������øa ����ʱ����ͪ����������塣�ڱ�ͪ������øϵ�İ����£��������dz�������������øϵ�İ����£��������dz�������øA��ͬʱ����1molATP��1molNADH��ά����B1B2��������������μӡ���Ӧ�Dz�����ġ�
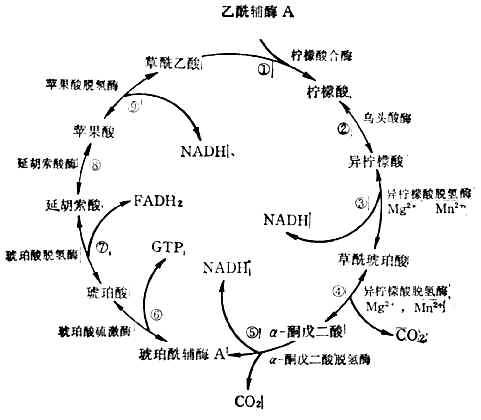
���� ��3��������ѭ��������øA�ϵ��������������ǵ�̼ԭ�ӣ���������ѭ������2molCO2����������ǵ���������Ӧ���������н��С���һ���̷�9����ͼ4-12����
����������ѭ���в���3molNADH,1molFADH2��1molGTP��
������NADH����ͨ��FAD��ϸ��ɫ����������ͬʱ�ų�������ÿmolNADHͨ����һϵ�з�Ӧ���ų���������220kJ��52.6kCal������Щ������ATP����ʽ����������ÿmolATP�ܲ���30.5Kj(7.3kCal)��������ÿmolNADHRNUD���������������������ϳ�7molATP������ָ�ڱ�״����25�棬1������ѹ��������Ͳ���Ũ�ȶ���1M/LPH7����˵�ġ���ģ�����ڵ�����ʱ��ʵ��֤��ֻ�ܺϳ�3molATP�����÷�Ӧʽ��ʾ��
����NADH+H++1/2O2 NAD++H2O+����
NAD++H2O+����
3ADP+3H3PO4+����3ATP+3H2o
���� ǰ��һ����������Ӧ������һ�������ữ������ż����һ�𣬳�Ϊ�������ữ��
ͼ4-11���ǵĽͽ�
����ʵ��Ҳ֤��FADH2����ʱ����2��ATP��GTP������1molATP��������Щ��ֵ����������ȫ����ʱ�������õ������ɹ������£�
�����ٽͽ⣺�����ǡ�2��ͪ��+8ATp
�����ڱ�ͪ��������������øA��
���� 2��ͪ���2������øA+2CO2+6ATp
������������ѭ����
����2������øA��4CO2+24ATp
ͼ4-12������ѭ��
������ÿmol������������������6molCO2ʱ�ɲ���38molATP������������õ�������38��30��1140kJ[��38��7.3����277kCal]��ÿmol������������6molCO2��ˮʱ�������ܵĸı���2870KJ��686kCal�������ԣ���еЧ��=1140/2870��10=40%��
�����������ñ�״�������ּ���ģ������ڸ�һЩ��Լ60%��
������������ɵ�����Ҳ�ɿ���̼ˮ�����ṩ�����������������������У���������ѭ����ࡣ֬���͵��������Ҳͨ��������ѭ��������ʳ��������ṩ��������90%����������ѭ����
���������Ǵ�л�����һ��;������ͨ�����Ƕ����ᣬ��������һ����ͨ·�����ڲ����˘y���ֽ�������-����ͨ·��
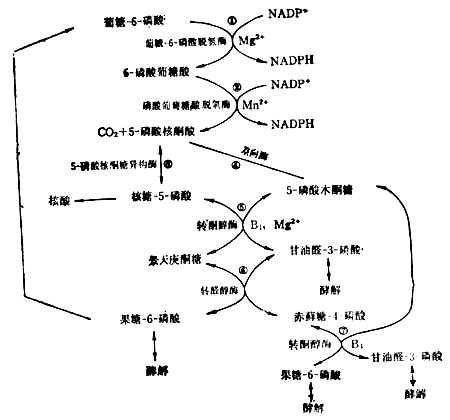
������4������-����ͨ·��һͨ·Ŀ�����ṩ�ϳɺ�������ĺ��ǣ��Լ��ϳ�֬���������̴��������������Ӧ��NADPH����ԭ��ø��ͼ4-13����������Լ��30%�������Ǿ�����һͨ����л����ϸ������һͨ·Ҳ�ܻ�Ծ������ʧ���ɵ�����Ѫ��ƶѪ����Ӧ����ϸ�����н��С�
��������-5-����ȿ����ں��������ϳɣ��ֿ���5-����ľͪ�Ƿ�Ӧ��������ȩ3-�������-6-���ᡣ��������һ��·��Ҫ��ϸ��������NADPH�ͺ��ǵ���Ҫ������������ϸ��NADPHҪ�ö࣬��ʣ�����ǿɾ�����ȩ-3-�������-6-����ͽ⣬���ɹ���-6-����ת���������-6-�����������һͨ·��
������һͨ·��ά����B1�μӡ�Ҫʹ��·��ͨ����ʳ������������ά����B1��5-�����ͪ��ת��Ϊ������-6-����ķ�Ӧ������������ά����B1�μӡ�Ҫʹ��·��ͨ����ʳ������������ά����B1��Ӫ��״����
������5����ԭ�����ǵ�����ϸ�����������Ժ���Ŀɴ�������������ϸ������ѹ�����ߣ�ϸ���������Ǻϳɴ���ӵ���ԭ����Ҫ����ʱ��ԭ�ٷֽ������ǡ������ԭ�����������ǣ�����ά��Ѫ�ǵĺ㶨������ԭ�����������ǣ�ֱ�����ڹ���������
ͼ4-13������-����ͨ·
����4��2��3����Ӫ����л�Ĺ�ϵ
������ϰ�������ͣ������Ϳո��������������Ҫ�߱��ܴ���̼ˮ���������ȱ��ʱ��֤������Ӧ�����ױ��졣
�����ͺ�Ѫ�����ظߣ��ȵ����������ӣ�����Ѫ�������ڼ��١�ʹ����������ǽ�����ࡢ�����֬����֯����ǿø�������������ѵ���������ԭ������ԭ�ĺϳɡ�������ԭ�Ĵ���������ɰ������Ǿ�������DZ�ͪ��ԭ�ɸ���-3-���ᣬ��������øA�ϳɵ�֬���ᣬ����NADPH�ṩ������������֬�����ٺ͵����ʽ�ϳɼ����ܶ�֬������Ѫ�����͵�֬����֯���档�����̼ˮ����Ҳ�ܺϳ�ijЩ�DZ��谱���ᡣ
�����ͺ�Լ4h����������ʳ���е������ǡ��ȸ�Ѫ���ط������ӣ��ȵ��ؼ��٣�תΪ���ø���ԭ�Թ���Ѫ�ǡ�����ԭ���������࣬�����ܶ��õIJ���70g,����һ�����ϵ����ġ����ԣ��Ӳͺ�8h��ʼ����������֬�����ṩ���������Ϳɱ���ǣ�����֬������ռ�������ࡣ֬����ֻ�ܹ��ܣ�����ת��Ϊ�����ǡ���ˣ�Ѫ�ǵ���Դ���ò�ת�����ˡ�һ��˵��������������������⣬���������ᶼ���ܳ��ǵģ����������Ĺ۵㿴��ֻ�б�������Ȱ������������ǵ���Դ����ʱѪ�ǵ�90%���Ա����ᣬ10%���ԹȰ��ᡣ
���������ܰѼ����������ı��������óɱ�ͪ������������ǡ��ո�ʱ�������н���ѪҺ�İ��������ӣ���������30��40%�DZ����ᡣ�����⵰���ʵı����Ậ������10%������ı�������������ģ������Ǿ��ͽ������ͪ�ᣬ�پ�ת�����ñ����ᡣ�����������ǵ�����û�����ӡ����ӵı�����ʵ���������������ᡣ
��������ѹȰ���ȥ����æ�ͪ����ᣬ�ٺ������ij��ǰ������������ת�����ö��ñ�ͪ�ᣬ���������ǡ���-ͪ�����Ҳ�ܾ�������ѭ���γ�ƻ���ᣬ���������ǡ�