2��������
2.1������
�����ʼ��ǹ�����֯��ϸ���Ļ������ϣ����������ʽ��������������������ٴ��ϣ������ķ�����Ԥ�������ƺ�Ԥ��Ҳ�������ڵ����ʵ�Ӫ��״��ϢϢ��ء�
Ŀǰ�����ʵ��о��ڻ�ѧ�ṹ���ռ乹�����﹦�ܼ�����ڿ��Ƶȷ��涼�кܴ�չ��ֻ���ڵ����ʵ��������������⵰���ʵ�Ӫ�����壬���ܸ��õ����ٴ���Ӧ�á�
2.2�������ʵ�����
2.2.1�������ʵĻ�ѧ
�����ʷ������������ӣ�������Լ��5��000���������������λ�ǰ����ᣬͨʽΪ��
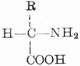
���ڰ������Ȼ����ڦ�-C�ϣ��ʳ�Ϊ��-�����ᡣʽ��R��ʾ������������ͬ�������������Ͳ�ͬ����ɵ����ʵİ�����һ����20�֣��������ᰴһ��������˳�����ļ���������������ɳ������ļ�����һ��������л��Ħ�-�Ȼ�����һ��������л��Ħ�-������ɣ�������ʾ��
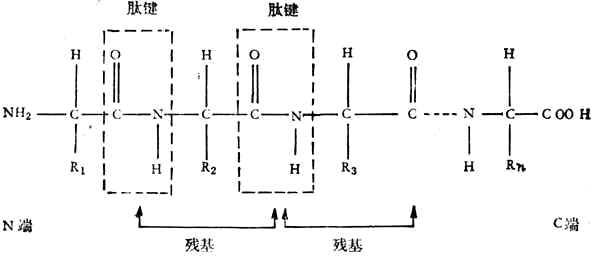
һ�������ʷ�����һ����������ɣ�ÿ������Լ����20�����ٸ�������л��������İ����˳�ΪN-�ˡ��Ȼ��˳�ΪC-�ˡ���������һ���ṹ�������ṹ�������ṹ���ļ��ṹ��һ���ṹָֻ�����а��������е�˳�����ļ��ṹ�������������ռ����еĹ�ϵ������������1��000��000�ĵ����ʶ����ļ��ṹ��
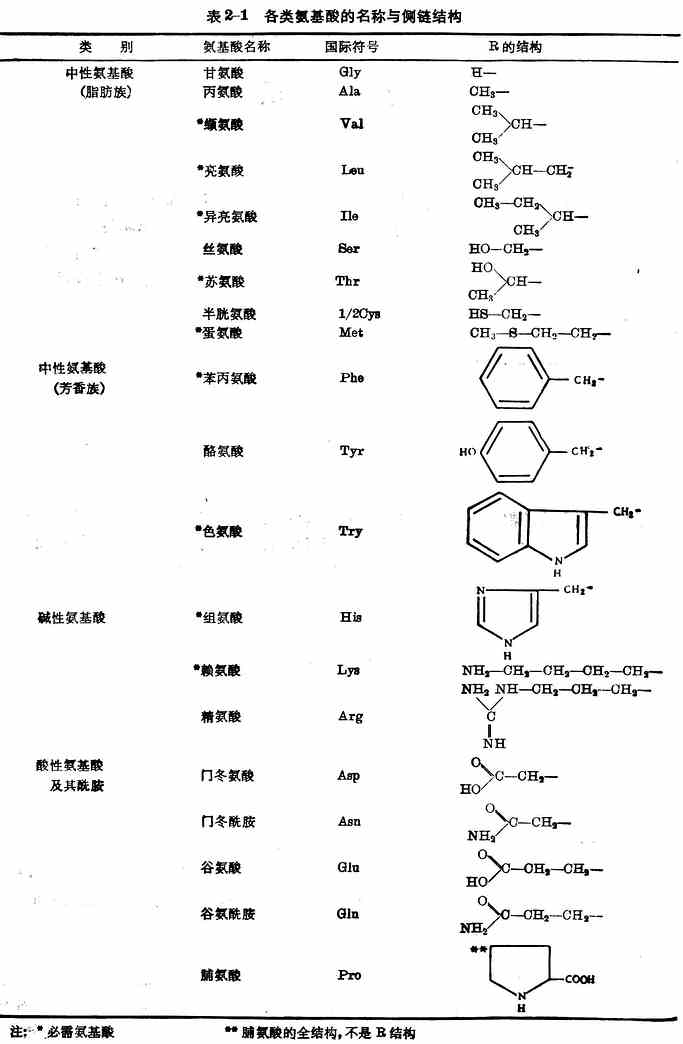
���հ�����������������Ȼ����������ɽ�20�ְ������Ϊ���ࣨ��2-1����
��20�ְ������������ܲ��뵰���ʺϳɣ���������������ȱ�ٵġ������������ᡢ�����ᡢ�����ᡢ�����ᡢ�������ᡢ�հ��ᡢɫ���ᡢ�Ӱ��ᡢ�鰱�ᣨ��2-1��9�֣������ڲ������кϳɣ���ϳ����ʲ������������Ҫ��������ʳ�﹩������Щ�������Ϊ���谱���ᡣ9���е��鰱����Ӥ�����谱���ᣬӤ��ȱ��ʱ��ʪ�����11�������������кϳɣ���Ϊ�DZ��谱���ᡣ���ǰ��װ�����Ұ����������ֱܷ��ɵ�����ͱ�������ϳɣ������ְ������������ʳ�к����ḻ�����н�ʡ�������뱽���������ֱ��谱��������ã������ʱ��Ϊ���谱���ᡣ
ע��*���谱���� **�������ȫ�ṹ������R�ṹ
�����ʺϳɺ�����ijЩ���������������������װ���ɽ���������γ�һ���װ����ԭ�е���������ǻ�����Ϊ����������⵰���е��鰱�ᾭ�����ϳ�3-���鰱��ȡ���Щ��������߱���й������ֽ��л�������ٺϳɵ����ʡ�
�Ӱ�����Ľṹ�������ʰ����⣬���ఱ����Ħ�-̼ԭ�Ӷ��Dz��ԳƵģ���������������ԡ�����-̼ԭ�ӵĹ��ͺ���ȩ�Ƚϣ����Լ����������еİ����ᶼ��L�͵ġ�L�ͺ�D�Ͱ�����Ĵ�л��Ӧ��ͬ���������������е�L�Ͱ��������D�͵Ľ����ڵ�����ͱ������ᡣ
����������Ϻ��а������Ȼ����ʳ����Ե���ʣ�ÿһ�ְ����ᶼ���ض��ĵȵ�㡣����������������ӻ��������Ρ��봼�����������л����γ��������������������ϻ����в���������ţ������������ʽ��л�ѧ��Ӧ����������л���á������������У�����������ijЩ�Լ�������ɫ��������ɰ����ᶨ�ԡ������ⶨ�Ļ�����������������������ͪ��Ӧ��������ɫ�����
�������ɰ�������ɣ��������백�������ƵĻ�ѧ�����⣬���н������ʺ����̡����Ե��������ʿ������ұ���������༫�Ի��ţ�����Һ���и߶ȵ���ˮ�ԡ���˵�����ˮ��Һ��һ�ֽ��ȶ�����ˮ���塣��������ˮ��ȫ���Ժʹ���ˮ�ֽ�ϣ��γɸ��������Բ�ͬ�Ľ���ϵͳ������γ��˸�����֯ϸ����ͬ����״�����Ժ�ճ�ȡ�
�����ʵķ�����ܶࡣΪ�˴Ӳ�ͬ�Ƕ��˽⵰���ʵĸſ����ֽ����ַ����������¡�
���յ������б��谱����ĺ������ࣺ
��1����ȫ���� ����������к���ȫ����������谱���ᣬ��ᵰ�ס��ѵ��ס�
��2������ȫ���� �����������ȱ��һ�ֻ���������谱���ᣬ���������
���յ����ʵĽṹ���ܽ�ȷ��ࣺ
��1������ ��������ֲ����֯�еİ��ס���ֲ�����еĹȵ��ס����ܵ��ף����ж�����֯�к���������Ƚ϶���㾫���ס��鵰�ס�
��2��Ӳ���� �����ܽ����͡�����������ë����ָ�ס��㡢���еĽǵ���Ƥ�����Ǹ��еĽ�ԭ���ס����Ե��ס�
��3����ϵ��� �����ڵ�������������ɵ����ס���֬������֬��ɵ�֬�����ڹǸ졢���졢����Һ�����ǽ�ϵ�ճ���ס��ǵ��ף�����ᡢѪ���ء�������ϵĺ˵��ס�Ѫ�쵰�ס��������ȡ�
���յ����ʵĹ��ܷ��ࣺ
��1�����Ե��� �����������������һ���л��Եĵ����ʣ���ø�����ص��ס����ͺʹ��浰�ס��������ס����嵰�ȡ�
��2���ǻ��Ե��� ���������Եġ��������ﱣ����֧�����õĵ����ʣ��罺ԭ���ǵ��ס����Ե��ȡ�
���ϸ��ַ��������һ���ľ����ԣ����ſ�ѧ�о��ķ�չ�������ϲ���������
2.2.2�����백�����л��
���ڰ������н���ͺ����������֡�����͵�Ũ��ƽ��Ϊ2mol����������Լ0.01mol,Ϊ����͵�0.5%�������������֯�а�����ķֲ������������ͱ��谱��������֯��Ѫ���е�Ũ�ȶ��ܵͣ��������ᡢ�Ȱ��ᡢ�Ȱ������ʰ������ַDZ��谱��������֯�нϸߡ�Ѫ������ͣ���2-2����˵���⼸�ַDZ��谱�����ܹ㷺��ϸ���ںϳɺʹ�����������������谱�����Ũ�������ÿ�հ�������Ҫ���Ƚϣ���2-2�������백�����ÿ�ձ���ת�����β������������������Ҫ����
��2-2 ����50g�����ڰ�����Ũ�Ⱥͱ��谱������Ҫ��
| ������ | ||
| g��100g-1ʳ��* | KJ��100kJ-1ʳ��** | |
| ƻ�� | 0.3 | 2.8 |
| ���ף��ϰ��� | 6.7 | 7.8 |
| ���� | 18.1 | 52.1 |
| С��ۣ���ǿ�ۣ� | 9.4 | 10.7 |
| ���� | 2.3 | 11.9 |
| ������ | 26.2 | 19.2 |
| ������ | 16.7 | 20.2 |
| ���� | 14.7 | 34.6 |
| �ƶ� | 36.3 | 35.2 |
| ���������� | 7.4 | 41.1 |
| ţ�� | 20.1 | 46.7 |
Ѫ�����백���ᾭϸ��Ĥ������֯ϸ������ϸ����Ĥ�Ͼ���ת�����ԡ����ԡ����Ը�����𰱻�������壬�����в�ͬ������֮���������������á�
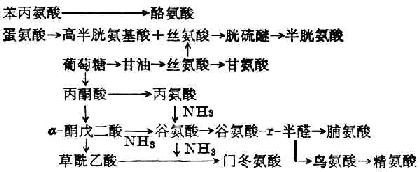
�����������ڴ�л;�����Թ���Ϊ���֣��ٲ�����֯���ס�����һ��ʱ���������֯���ķֽ⣬���������백����⣻�ڽ��зֽ��л����̼���γ�CO2��������ת��Ϊ��ԭ��֬��������䰱���γ������ų����ۺϳ�������������������ʼ���ᡢ�������ء���Щ���ʼ������ⲻ�ٷ������백����⡣���⣬���ϳ������DZ��谱���ᡣ
2.2.3��������ת��Ϊ������������
��������������Ҫ�������ϳɵ����ʣ��������ںϳ�����һЩ���������Ե����ʡ�
�ϳɷDZ��谱����Ұ���Ͱ��װ���ֱ��ɱ�������͵��������������������DZ��谱�������������ѭ���������Ħ�-ͪ������������������ͪ���γɣ�ͼ2-1����
�����ϳɲ��ձ�����ڸ�����֯���籽������ֻ�ڸ������ǻ�ø�����γ��Ұ��ᡣ
ͼ2-1 �DZ��谱����ĺϳ�
���ʺ���व�����ϳɣ����ʺ���़�ɴ�ʳ�﹩����Ҳ�����������кϳɡ��ϳɵ���Ҫԭ�����Ŷ����ᡢ�Ȱ��������ʰ���ȡ�
����ͼ�����������ϳɣ�������ʳ�﹩����Ҳ���������ڴӵ����ᡢ�ʰ���;�����ϳɡ�������ѪҺ�������⣬���������գ�ÿ��ת����Ϊ2%�����ڴּ����Լ�������ἡ�����ʽ���ڹǸ켡���ھ�ֹ�ļ������������ἡ��Ϊ��������ƣ�͵ļ����У����ἡ��Ũ��ȴ�ܵ͡������������ἡ�������ἡ��ת��ø�������£�ת��Ϊ�����ATP�Ľ����

�����Ӧʹ���������������£��ܴ����ἡ���ø��ӵĵ���������ATP�����ۼ���������ἡ����Ҫͨ����ø��ˮ��Ӧ��������������γɼ��������������������ã�����ѪҺ�����ų���ÿ�ռ��������ɵ����൱�㶨��ԼΪ�ܼ�����1.7%��
2.2.4�������ʵ�����ϳɺ���ת
�������и߶ȵ������ԡ�ʳ�ﵰ�ױ��뾭������ˮ���Ϊ����������գ��ٺϳ���������Ҫ�ĸ��ֵ����ʡ������ʵĺϳ���ϵ��Ҫ����ʹ���Ǻ��ᣨmRNA����ת�˺��Ǻ��ᣨtRNA�������Ǻ˵�������ᣨrRNA����ijЩ���������ӹ�ͬ��ɡ�mRNA�ǵ����ʺϳɵ�ģ�壬tRNA�ǰ��˰����ᣨԭ�ϣ��Ĺ��ߣ�rRNA�൱��װ������ٽ�����������ļ����ϡ����ְ������ڸ��Եİ��˹���Я���£���װ����ϰ���ģ���Ҫ���д�������ϣ����ɾ���һ������������˳����ض����������ϳɺ�Ķ��������еľ���һ���������е����������������ǡ�֬�ʵȽ�Ϻ��γɾ���������Եĵ����ʡ����ϳ�ԭ�ϣ��ر��DZ��谱���ᣩ��������ʱ��������ϸ���ڵ����ʺϳɼ�����ֹͣ��
ÿ�յ����ʺϳɵ���ȡ�����������ϳɸ���ø������֯ϸ������Ҫ��������֯ϸ���ϳ���ֽ�����ʲ���ܴ���С��ճĤÿ1��2�����һ�ζ���ϸ����������ԼΪ120�죻������ͬλ���о�����ÿ���ܺϳ�Ѫ������10g����ά����2g�����������Ǹ��н�ԭ�����������ȴ����300�졣�ø���������15N�����᷽�����Ʋ�ͬ������ɵ����ʺϳɵ����ʣ��ó�����Լ70kg�ij�������ÿ�յ����ʺϳ�������200g����2-3�������������������ÿ�յ�������Ҫ�������������ɱ��������������¾ɵ����ʷֽ�����İ������������ڵ����ʵ��ٺϳɡ�
��2-3 ��ͬ�������ܵ����ʺϳ���
| ������ | ���� | ���أ�kg�� | ���䣨��Χ�� | �ܵ����ʺϳ��ʣ�g.kg-1.d-1�� |
| ������������� | 10 | 1.94 | 1��46�� | 17.46 |
| Ӥ�� | 4 | 9.0 | 10��20�� | 6.9 |
| ���� | 4 | 71 | 20��23�� | 3.0 |
| ���� | 4 | 56 | 69��91�� | 1.9 |
2.2.5��������Ľ���
���ְ����ᰴ���ض��Ļ�ѧ��Ӧ���н��⡣ͼ2-2�����������ڰ����ή�����Ҫ�����Ͳ��

ͼ2-2 ��������Ҫ���ⳡ�������
��7�ֱ��谱������Ҫ�ڸ��ཱུ�⣬�����������ᡢ�����ᡢ�Ӱ���3�ֱ��谱���ᣨ֧�������ᣩ��Ҫ�ڼ������Լ��������н��⡣֧���������ڼ����о�ת�������ñ�Ϊ��ͪ��Ȱ��ᣬ�̶��ֱ��γɱ�����Ȱ��������پ�Ѫѭ���ֱ��͵�����ͳ����ڳ���ת��Ϊ������Ȱ��ᡣ�����ᾭ�ž���������࣬�䰱���γ����ء�̼�ܾ���ԭ��������תΪ�ǣ�ͼ2-3����
���ص��γɼ���ȫ�������ڸ����У��������Ψһ���о�����ø����֯�������ǰ���CO2�ϳɰ��������ᣬȻ���ϰ��ᡢ����������������ᡢ�������һϵ�кϳɷ�Ӧ��ͼ2-4����������ھ�����ø�Ĵ��·ֽ�Ϊ���غ����ᡣ�����ط����غϳɵ�ѭ�������������ų���
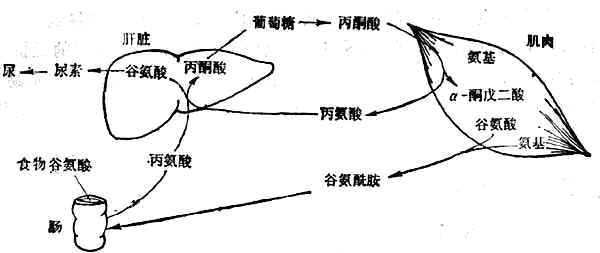
ͼ2-3��������-������ѭ��
CO2+NH3+ATP
��
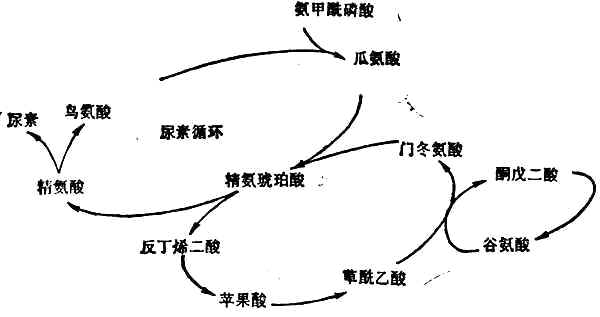
ͼ2-4 ���غϳɵ�;��
���ǵ����ʴ�л�����ղ���֮һ��������ϳɡ���Ѫ�������ĹȰ�������������Ľ���С����Ƥϸ���У����Ȱ�����ø��ˮ��Ϊ���Ȱ����Ρ��Ȱ������ڹȰ�������ø�Ĵ��£��ֲ������ͦ�-ͪ����ᡣ�������ų�����-ͪ����ᾭ��ԭ���������γ������ǣ�����Ѫѭ���������á��ڼ��������ȷ������ж�������£������࣬�������й���ֳ��˹��������ǵij����������γ�Ҳʹ���屣���������ӣ����������ӽ������к�������ų���
2.3�������ʵ���������
�������ǹ���������֯����Ҫ�ɷ֣��ǹ�������Ψһ��Դ���京��Լռ�����ܹ�������45%���������һ��ϸ����֯���ɵ�������ɡ����������Ҫ�������õ����ʣ�ȱ�ٵ����ʾͲ����ڡ����д����õ�ø�����ڸ��ִ�л���̵ĵ����أ�������С���ӡ����ӡ����ӵ����䵰�ף����������ļ������ף��з������ܵ������ף����ɻ���֧�ܵĽ�ԭ���ȡ�
��һ������¹������ܲ��ǵ����ʵ���Ҫ���á���������֯ϸ�����ϸ��¹����У������ʷֽ�ɰ��������һС���ֲ������ö��ֽ���ȣ�Ҳ��һ�������յİ����ᣬ������ʳ��������嵰�ϳɵ���Ҫ�����������ȡ�����ÿ���������ܴ�Լ��10��15%���Ե����ʡ�����������£����Ǻ�֬�����벻��ʱ�����������ڲ������ܡ�
���崢�浰���ʵ������٣���Ӫ������ʱ��Ҳ����ֻ���嵰��������1%���ҡ����ֵ����ʳ�Ϊ�����ף���Ҫ���ڸ��ࡢ��ճĤ�����٣���ʧ������ٹ���û�иı䡣����ʳ����ȱ��ʱ����֯���ֽ�졢�ϳ�������������һϵ�������������ı���ٴ����֣���ճĤ�������ٽ����ۼ����ٴ�����Ϊ�������ղ�������к�������ά�������ṹ�빦�ܣ�����֬������Ѫ�����ϳɷ����ϰ���ø�Ļ��Խ��ͣ���Ҫ�ǻ���������ø�Ȱ�������ø���ͣ����ڼ��⵰�ϳɲ�������ּ���ή��������ϳɼ��٣��Դ�Ⱦ���ĵֿ����½�������������Ƥ�ʹ��ܼ��ˣ����ѿ˷�Ӧ��״̬����ԭ�ϳ�Ҳ�ᷢ���ϰ���ʹ�˿ڲ������ϣ���ͯʱ�ڿɼ������������������������ϰ��������ʳ������벻�㣬�����γ�Ӫ����ˮ�ף�����ʱ����������
2.4����ʳ�����ʵ�����
2.4.1�������ʵ�����������
ʳ�ﵰ����θҺ����ø�������£�����ˮ�⣬��С������������������չ��̡�θ����øԭ��θ�ײ������Ų�����ϸ�����ڣ���θ����Ѵ��ڵ�θ����ø�����£��ͳ�һ���ֶ��ģ��γɾ��л��Ե�θ����ø��θ����ø�����ý�����רһ�Խϲ��ճҺ�����⣬ֻ�ܴٽ�����ˮ���Ե�����ˮ���Ϊ���ģ���Ҫˮ�ⱽ�����ᡢ�Ұ������������ɵ��ļ���
��Һ�����ȵ���ø����Һ�鵰��ø�����Ե���ø������ø���Ȼ���øA��B������ø����ø��������ˮ������ú�רһ�Զ���ǿ������θ�г������ˣ�ʳ�ﵰ��δ��θ����ø�����ã��������ʲ����ܵ����ص�Ӱ�졣�ȵ���ø�������м�������л����Ȼ���ɵ��ļ��������鵰��ø��Ҫ�������з����就����л����Ȼ���ɵ��ļ������Ե���ø��������֬���就����л����Ȼ���ɵ��ļ����Ȼ���øA��������������л�C�˵��ļ����Ȼ���øB�����ڼ��������л���C�˵��ļ���ͼ2-5����
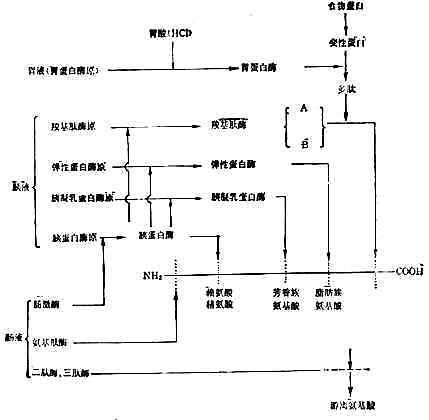
ͼ2-5 θ�����ڵ����ʵ�����������Gilter�ģ�
�����ڵĸ�����ø��������øԭ������ʮ��ָ����ű���������á��ȵ���øԭ��С����Ƥϸ��ˢ״Ե����ij���ø���ʧN�����ı���ȵ���ø���ȵ���ø�ټ��������鵰��øԭ�����Ե���øԭ���Ȼ���øԭ��ʹ�����Ӧ��ø��ͼ2-5�����ȵ���øԭ�ķ����ܳ���ʳ�ﵰ��Ӱ�죬���ȵ���ø��ʳ�ﵰ�����Ϻ�����ȵ���ø�����������ٷ��ڡ�
��øˮ�⵰�IJ����1/3Ϊ�����ᣬ����Ϊ���ģ�10�����������µ�����������Һ����ø���٣����ڳ�ճĤϸ����ˢ״Ե�Ͱ�Һ�У��ֱ��ж��ֹ���ø���ܴ�������N����ˮ����������֮Ϊ������ø��ˢ״Ե���е�ø��ˮ��2��������������ɵ��ģ���Һ�е�ø��Ҫˮ����ġ����ġ�һ����Ϊ��������£��������ϵ���������ˢ״Ե����ˮ��Ϊ���Ļ���ģ�������ϸ�����ٽ�һ���ֽ�Ϊ�����ᡣ
L-������Ͷ���������Ҫͨ��������������ת�˴ӳ������ա��ڳ�ճĤϸ��Ĥ�ϣ��������ղ�ͬ���ఱ����Ͷ������ĵĸ߶�רһ�Ե�ת����ϵ��D-�������������Ӧ�ĵ�L-����������ͬһת��ϵͳ��������ϵͳ������Զ����L-�����ᡣ�ڵ����ʴ�л����������У��������ĵ��������ʱȰ�����죬���Ժͽ�Ϊ��ˮ�İ���������������ֱȼ��Ժͽ�Ϊ��ˮ�İ�����Ϊ�졣
�����еĵ����ʲ���ȫ��������ʳ��������������ճĤ����Ƥϸ��������ø����Դ�Ե����ʡ��ݹ��Ƴ���ÿ������Դ�Ե���35��130g����������������ʱ��Щ���������ڵ����ʴ�л����Դ�Ե��ײ��������θ������ã����ұ��汻��Һճ���ױ������ʽ�������������Դ�Ե����������س����ܳ���������ա��ڻس�ĩ�κʹ���ϸ����øҲ�ֽܷ⵰���ʣ����˵Ľ᳦�������հ����ᣬ���ɵİ�����ֻ�������ų���
2.4.2�����������
��ʳ�����ʺ��ž���Ѫ�а������Ũ�Ⱥ�ģʽ���иı䣬����ѭ��Ѫ�а�����ı仯�����������ڸ����DZ��谱���ᣨ֧����������⣩���зֽ��л����Ҫ�����������ܿ�������İ����ᣬ���ܰ��ջ������Ҫ�������л���ʡ�
������ʳ���������İ�������ɸ��ཱུ��Ϊ���أ�С�����ڸ������ںϳɵ����ʡ���Elwyn(1970)����������ʳ��140g��������İ�����Լ57%����Ϊ���أ�14%�ϳɸ��൰�ף�6%�ϳ�Ѫ�����ף�����23%������ѭ����
���������������ʱ������ͨ��ø�����ü��Կ��ơ���������£������зֽⰱ�����ø������Խ��٣���������������ʱ��ø�ĺ�����Ӧ�Ե����ӣ�ʹ����������Ҫ���İ�����ֽ��ƻ���
�����е����ʵĺϳ�����ȡ���ڰ������������Ͱ������ģʽ����ɴӺ˵�����ĺϳɷ�ӳ�����������������������ٻ�ƽ��ʱ����˵�������Ϊ�ͺ˵�����͵��˵����壬���Ǻ���ֽ⣻����������������ͱ�������ʱ���෴��
2.4.3��ѪҺ������Ũ�ȵĵ���
Ѫ�����백�����ǰ������ڸ���֯��ת�˵���Ҫ��ʽ������ͨ��ά��Ѫ���������Ũ�ȣ�������ͬ����������֯��Ҫ��֮���ƽ�⡣��������Ѫ�����백������Ũ��Ϊ350��650mg��L-1����ҹ���������Լ30%�����𰱻�����ҹ��һ�㲻����50%��
Ѫ�����������ܸ�����ڣ�����������������������л��ʱ��Ѫ���������Ũ�Ȼἱ����������ֱ���������ͳ�������Ժ���ͬ��ɫ��������ϣ���ɫ���Ậ������������ά����Ҫ���������������ɫ������Ҫ��ռ����0.1%��������ɫ������Ҫ��ռ����0.03%��ʱ��Ѫ��ɫ����Ũ�����ߡ����ַ��������ڲⶨ������백�������Ҫ�������ó���ɫ�������Ҫ��Ϊ3mg/kg���ء������õ�ƽ�ⷨ�ó�����Ҫ�����һ�¡�Ȼ���������������ᣨ�������ᣩ�IJⶨû�еõ�����Ľ����
Ѫ��������Ũ��Ҳ����ʳ�ǵ�Ӱ�졣�ڽ�ʳ�Ǻ��ȵ��صķ������ࡣ���ȵ��ص������£��������������뼡�⣬��ʹѪ���е�Ũ���½���������֧����������½���Ϊ���ԣ��ڽ�ʳһ�������Ǻ�ɽ���40%����ijЩ�����ᣨ��ɫ���ᣩ��Ӱ����١�
�����������ϸ��������Ҳ������Ѫ�������������������Ũ�ȡ�������ɫ�������������ɫ����ĺ������ߣ�5-��ɫ�����ࣻ�������ʳ�ǣ�����Ѫ����֧�������������½����ٽ�ɫ�����������֯��ͬ��ʹ����5-��ɫ�����ߡ�
�������ò�����5-��ɫ���Ĵ�л�йأ������ڲ�������£����Թ��ܵ�Ӱ��Ҳ����Ҫ���塣����Ӳ�������ܲ�ȫʱ���ڳ���ϸ�������²����İ��Ͱ��������������ڸ���ֽ���ת�䣬������ѭ��ͨ��Ѫ�����Ͻ�����ϸ�����籽��������ϸ�������±�ɱ��Ұ���������ת��Ϊ��-���Ұ�������������Ϊ�������Ӱ������ļ����ʡ�����������£��ܱ�������ڵİ����ᣨ��ɫ���ᡢ�������ᣩ����ʱҲ�����ܿ��ƣ�����Ѫ����������֯�����⣬����������£���Ҫ�ڸ��ཱུ����ȵ���Ҳ���ٷֽ⡣Ѫ���ȵ��غ������ߣ��ٽ�֧����������뼡�⣻Ѫ��֧���������½�����������ɫ�����������ϸ���ı�������������������ʹ����֯��5-��ɫ���������࣬����λ��ԡ����������۵㣬���λ��Ի����Ժ���֧�����������ʳ����Һ������Ч�ؽ��֢״��
2.4.4��������������
�Ǹ켡������������֯��Լռ���ص�45%����ˣ��ڵ����ʴ�л�У����ⰱ����Ĵ�лռ�൱��Ҫ��λ�á������ǰ������л�ij�����Ҳ��֧���������л����Ҫ�������ڼ��ⰱ�����л�У��ȵ�������Ҫ�ĵ������á����ܴٽ������ᣬ�ر���֧�������ᣬ���뼡����֯���ٽ����⵰�ĺϳɣ���������ֽ⡣������Ƥ�ʼ��غ��ȵ��ص��������෴��
�ⶨ���⵰�״�л�ķ�����Ҫ�����֣����Ƚϼ������Ѫ�а����Ậ���IJ��Ͳⶨ����3-���鰱����ų�����
��ʳ�������ͷŴ���������Ȱ������������൱��70kg���ص�����ÿ�ն�ʧ75g���⵰�ס����������������Ƿֽ�����ı�ͪ���백����ֽ��ͳ��İ����ϳɵģ�ͨ��ѪҺ������࣬�ڸ��ྭ��ԭ�������γ������Ǻ����ء�ǰ�߷��ؼ��⣬�������ã��γ���������-������ѭ�������������ų���ͼ2-3�����ǰ�����ֽ�����İ�����ѪҺ���������ʽ���Ȱ��᰷��Ҫ���볦����֯�����࣬�ڳ�������ת��Ϊ��������������ࣻ�������зֽ��ͳ�������Ϊ����Ҫ��Դ��ͬʱҲ������ԭ�������á�
3-���鰱���Ǽ����м������ס����ֽ�ʱ��3-���鰱����ų����������о�ʳ��ɷֶԼ��⵰�ֽ����ʵ�Ӱ�졣�����������ͨ�ġ�ȱ�������ʺ�ȱ�������뵰���ʵ��������ϡ����ֵ�����ȱ���飬����3-���鰱����ų����ܿ����ԭ�ų�����1/5�����Ե����ʺ��ų������ߣ������뵰����ȱ���飬����3-�������Ὺʼ���࣬������½���ʵ�������������ȱ��ʱ�����⵰��ֹͣ�ֽ⣬���������뵰����ȱ��ʱ�����⵰���ȼ��ٷֽ⣬Ȼ�������ӡ��Ӫ�������Ķ�ͯ��3-���鰱����ų����ͣ�Ӫ������������ߡ������߳��ڽ�ʳ������3-���鰱���ų��������½���
����3-���鰱����ų���Ҳ������ͼ��ص�Ӱ�졣������ÿ�������ص��ų������ڳ����ˣ������˵��ų����ֱ��С�����Ϊ�͡�
2.4.5����ƽ��
���ڵ���л�����ղ�����Ҫ�����ų�����Һ�������Ƥм�к�����������������������ĵ���ë�������顢�¾�����Һ�ȶ�ʧ��������δ�����յĺ���������ӷ��ų���
������Ҫ�ĺ��������������ء���������ͼ������������浰���ʵ�������죨��2-4����
��2-4 ���˲�ͬ��ʳÿ�����ų�����g��
| ����Դ | �ߵ����� | �͵����� | ��ʳ | |
| ��1�� | ��2�� | |||
| �ܵ� | 16.80(100) | 3.60(100) | 10.51(100) | 8.71(100) |
| ���ص� | 14.70(87.5) | 2.20(61.7) | 8.96(85.1) | 6.62(75.4) |
| ���� | 0.49(3.0) | 0.42(11.3) | 0.40(3.8) | 1.05(12.0) |
| ���ᵪ | 0.18(1.1) | 0.09(2.5) | 0.12(1.1) | 0.17(1.9) |
| �������� | 0.58(3.6) | 0.60(17.2) | 0.44(4.2) | 0.39(4.4) |
| ������ | 0.85(4.9) | 0.27(7.3) | 0.59(5.6) | 0.54(6.1) |
ע��������Ϊ���ܵ���%
��ͨ��ʳʱ�����ص�ռ�ܵ���80%���ϣ��͵�����ʱ�����ص����ͣ�����ʱ���������ߡ����������ų����ƺ�����ʳ���ĺ����ء�
��֯���ķֽ��л�ͺϳɴ�л���ڶ�̬ƽ�⣬����ƽ����õ�ƽ���ʾ��
B=I��?/FONT>U+F+S��
��ƽ�⣨B�������뵪����I�����ų�����[��U�����൪��F����Ƥ����S��]�IJ�����������������ų��ĵ���������ȣ�B���ڻ�ӽ��㣨���뵪�ġ�5%���ڣ�����Ϊ��ƽ�⣻��ͯ����������ʱ�ڣ���һ���ֵ����������ڴ�����BΪ��������Ϊ����ƽ�⣻˥�ϡ����ݵļ�����ijЩ�����Լ������ų������������뵪����BΪ��������Ϊ��ƽ�⡣
�������������ų�������ʵ�鷽����á��ڲⶨ�У�����ʳ��δ��ȫ�������������ļ�������ƫ�ߣ�������Ʒ��ʧ����Բ��ƣ��ų����ļ�������ƫ�͡����������ܺͿ�ʹ���������ų������10��15%��
��ƽ����������������Ӱ�죬�����н�ʡ�����ʵ����á���1973��FAO/WHOר��ίԱ�ᱨ�棬�������ߵ�����0.57g��kg-1���أ������ܹ�������ԣʱ����������ƽ�⣻�������ܹ�������ά��ˮƽʱ�����ָ���ƽ�⡣��ƽ����������ء�غͪ��Ƥ����̴��ͼ�״���صȼ��ص�Ӱ�졣��Щ�����дٽ������ʺϳɵ����ã���ٽ������ʷֽ⡢���ƺϳɵ����á�
2.5�������ʺͰ��������Ҫ��
����Ե�������Ҫ�����о���������100�������ʷ���������ϵķ�չ������50���ǰ��Rose���˶�������ֱ��谱������Ҫ��������һϵ�еIJⶨ���Ժ���������Ϲ�������ټ�ר�����ۺ��������ʺͰ��������Ҫ�����йص��о������������ڽ����С�
2.5.1�������ʵ���Ҫ��������
Ӫ���ص���Ҫ����ά�����������������ܺͽ���������������������������������Ⱥ�о����������Ҫ����ȡ�����Ǹ�����Ҫ���ƶ��ġ���������Ҫ���IJⶨ������Ҫ����㷨����ƽ�ⷨ���֡�
Ҫ����㷨��factorial method�����òⶨ���趪ʧ����obligatory nitrogen loss����ȷ����������Ҫ���ķ������ˣ�����ڽ�ʳ�����ŵ�����������ʧ����Դ���������൪��Ƥ�����ȣ���Ϊ���趪ʧ����Ϊά�ֽ�����ÿ�ն�ʧ�ĵ�������貹�����Ӳ��������Եó������ʵ���Ҫ�����˴�������ͨ�Ÿ�Ϊ���ź����ų����������½���Լ8��14���ﵽ�ȶ������ֵ����ֵ����Դ����FAO/WHOר��ίԱ����1965�������Ҫ����㷨��1973�������ģ���ⶨֵ�͵����ʹ������ļ������2-5��
��2��5����Ҫ����㷨����������ӵ����ʹ�������FAO/WHO1973��
| ƽ�����趪ʧ����mg��kg-1��d-1�� | |
| �� | 37 |
| �൪ | 12 |
| Ƥ���� | 3 |
| ������ | 2 |
| �ܵ� | 54 |
| ����������ӣ�%�� | 30 |
| ���ѵ��������ӣ�%�� | 30 |
| ����������mg��kg-1��d-1�� | 91 |
| �����ʹ�������g��kg-1��d-1�� | 0.57 |
| �����ʹ�������g��70kg-1��d-1�� | 40 |
ƽ�����趪ʧ�������������������죩���Եõ�����97.5%��Ⱥ��Ҫ�Ĺ�������Ҳ�ɰ�������ѧ�������Ĺ��ɼ�������15%�Լ��㹩�������˴��������㣬54mg����30%��70mg��Ϊ������ʧ�����òο��������ñ������㵰������Ҫ���������ѵ����ڳ������ʵ���е�������Ϊ95��98%��1965��������Ϊ��Ȼ�ο����ף����Ժ������嵪ƽ��ʵ����֤���ѵ��������ʽ�70%���ҡ���ˣ���1973��ļ����У�ΪУ���ѵ��������ʵIJ��㣬��70mg��������30%��91mg����0.091g��kg-1���ص�����6.25,�õ����ʹ�����0.57g��kg-1���ء���Щ���������ַ����Ļ����ϼ����ģ�����������Ĺ���������������1980�꣩Ϊ0.8g��kg-1���أ��ձ���1980�꣩Ϊ1.18g��kg-1���ء�
��ƽ�ⷨ��nitrogen balance method�����ڿ�����ʳ����ͬ�������ʵ�����£�����ﵽά�ֵ�ƽ��ʱ�ĵ���������������Ϊ���嵰���ʵ���Ҫ�������ַ�����Ȼ���ϣ����Ҳⶨֵ�������ڵ����ʴ�����������������Ӱ�죬��Ŀǰ����������Ϊ�ⶨ���嵰������Ҫ����һ�ַ�����1963����ʸ���Sherman�ĵ�ƽ��ʵ�飬��������˵�������Ҫ��Ϊ1g/kg���ء����ɷ��ͳ�ѧ��Գ������ӽ��е�ƽ��ʵ�飬�����ܹ�����ԣ������£������ʵ���Ҫ����0.9�� 1.0g��kg-1���ط�Χ�ڡ��ҹ���ʳ��ֲ����ʳƷΪ����ֲ�ﵰ�������ֵ�Ե͡���ˣ�ÿ����ʳ�е��ʵĹ�����Ӧ��1.1g��kg-1���أ���������63kg����Ϊ70g(1988������ȫ��Ӫ��ѧ��������)����ͯʱ����Ҫ����ĵ������Ա�֤����������1������Ӥ�������ʵ���Ҫ��������ι����Ϊ2g��kg-1���أ�ţ��ι����3.5g��kg-1���أ����ι����4g��kg-1���ء�1���Ժ����٣�ֱ�������˵�1.1g��kg-1���ء�������Ϊ��֤ĸ���̥��������Ҫ���������4��6����ÿ�չ���������15g����7��9����ÿ������25g����ĸÿ��Ҳ����25g��
2.5.2�����谱������Ҫ��
������Ҫ�����ʣ�ȷ�е�˵����Ҫ�����еİ����ᣬ��˲ⶨ���������Ҫ���Ȳⶨ�����ʵ���Ҫ������ֱ�����塣
�о���������Ҫ���ķ����Ǹ�ʵ���������ʳȱ��ijһ�ְ����ʳ�Ȼ�䲻ͬ���ĸ��ְ����ᡣ���ﵽ��ƽ�⣨���ˣ���ٽ�������������ͯ��ʱ�����������������ְ��������Ҫ����Rose�����õ�ƽ�ⷨ�ó��������Ӹ��ְ��������Ҫ�����Ժ���ѧ���о��˲�ͬ���䡢�Ա����Ⱥ�ı��谱������Ҫ�����õ�ƽ�ⷨ�ó�����Ҫ��һ��ѡ��Χ�ϴ��ֽ�Rose���˲ⶨ����Ҫ��ƽ��ֵ���ڱ�2-6��Ŀǰ�ҹ��ݲ���FAO/WHO��1973�����ݣ���Ϊ�������ӱ��谱������Ҫ������
��2-6 ������谱����ƽ����Ҫ����mg��kg-1��d-1��
| Ӥ����Holt�� | ��ͯ10��12�꣨Nakagawa�� | �������ӣ�Rose�� | ����Ů�ӣ�Hegsted�� | ���ˣ�FAO/WHO 1973�� | |
| �鰱�� | 25 | - | - | - | - |
| �������� | 111(5.8) | 28(7.0) | 10(3.3) | 10(3.3) | 10(2.9) |
| ������ | 153(8.1) | 49(12.3) | 11(3.7) | 13(4.3) | 14(4.0) |
| ������ | 96(5.1) | 59(14.8) | 9(3.0) | 10(3.3) | 12(3.4) |
| ������+�װ��� | 50(2.6) | 27(6.8) | 14(4.7) | 13(4.3) | 13(3.7) |
| ��������+�Ұ��� | 90(4.7) | 27(6.8) | 14(4.7) | 13(4.3) | 14(4.0) |
| �հ��� | 66(3.5) | 34(8.5) | 6(2.0) | 7(2.3) | 7(2.0) |
| ɫ���� | 19(1.0) | 4(1.0) | 3(1.0) | 3(1.0) | 3.5(1.0) |
| �Ӱ��� | 95(5.0) | 33(8.3) | 14(4.7) | 11(3.7) | 10(2.9) |
| �ܼƣ���ȥ�鰱�ᣩ | 680 | 261 | 81 | 80 | 83.5 |
��������ֵ�Ǹ���ԭ����ɫ����Ϊ1�ļ���ֵ��
ժ��White,PL et(1974):"Total ParenteralNutrition",p.65
ǰ���ᵽ�װ�����Ұ��������ڿ��Էֱ��ɵ�����ͱ�����ϳɡ���������ַDZ��백����ɷֱ��ʡ������ͱ������ᣬ���װ���ɴ���30%�����ᡢ�Ұ���ɴ���50%�������ᡣ
���嵰���ʺͱ��谱�������Ҫ������kg���ؼƣ�������������������½��������谱�����½��ķ��ȸ���Щ������ÿ�������ر��谱�������Ҫ����ԼΪӤ����Ҫ����1/8��������������谱�����ƽ����Ҫ������30%�����Ϊ97.5%��Ⱥ����Ҫ�����ٺ���Ӧ������ĵ�������Ҫ���Ƚϣ��ֱ�ó����谱�������Ҫ��ռ��������Ҫ���ı�ֵ��Ӥ��Ϊ43%����ͯΪ36%������Ϊ19��20%��Ӥ������Ҫ���ȳ��˸ߵ������ǣ�Ӥ����������ά�ֵ���Ҫ����������Դ������ʧ�İ����ᣩ�⣬����������������Ҫ��
���ֱ��谱�������Ҫ�������㹻����Ҫ�����ı��������ģʽ��ǡ������Ϊ����ϸ�������ʵİ�������һ���ı�������ʳ�������ṩ�ĸ��ֱ��谱��������ֱ�����������ܳ��Ϊ���������á����ȱ�����е�һ�֣���tRNA�Ͳ����ܼ�ʱ�ؽ�����Ҫ�ĸ��ְ�����ȫ������rRNA������������ò���������ã������ʵĺϳ�Ҳ�Ͳ���˳�����С���2-6�����ڵ���ֵ�о��˸���ģʽ��ʵ��֤���������������ij�ְ����ᣬ���������ģʽ�нϴ��ƫ��ʱ�������������Զ������л�ϰ�����ֶ���֢״����ÿ������������2%�����ᣬ��ʹ���������ٻ����Ρ�Ƣ���ȷ��������Ա��ԣ�����ʴ�ȣ��������ṩ�����㣬Ҳ��������������������㣬����ɳ���֬���Σ�ɫ����㣬�������ȱ����
��Ӧָ�������ܺͷDZ��谱����Ĺ�Ӧ�����ԣ������ʹ��2-6���оٵı��谱��������ܹ�������幹����֯����Ӫ��Ч�ܡ�
2.5.3��Ӱ�쵰������Ҫ��������
��ʳ�����н�ʡ�����ʵ����ã����ܹ������㣬�����ʽ������������ܡ�Callowag��1974���ȷ��ֹ�����7��8g������������ʱ��������ʧ�ͼ��٣������ܹ����ﵽ12600kJʱ���͵õ�����ƽ�⡣�����Ӫ��֧��ʱ���о���������Ҫ����ʵ���У�������ע���ڸ�ʱ���͵õ�����ƽ�⡣��Ҫ����㹻�����ܡ��Ȼ����Ե�������Ҫ����Ӱ�죬�����һ�¡�Consolazio��������Ӧ����38.7���½����е������Ͷ�ʱ���ֱۺ�Һ������Ϊ241mg��L-1�������Ϊ�����Ȼ����½����е������Ͷ�ʱ�������ʵ���ȡ��Ӧ��һ���������Ļ���������13��14%����Ashworth�ⶨ6�����������Ӧ�ߵ�ȫ����Һ�����ֺ���Ũ�Ȳ��ߣ�Weinen�ⶨ6��̹ɣ��������ĺ�Һ�����ֺ���ÿ�����ʧ��ֻ��0.5��1g��Thaper�����ֵ�����Ũ������ʱ������������Խ��ͣ���ˣ�������Ϊ��ʹ���Ȼ����ູ������£�Ҳ�������ӵ����ʵĹ�������
�������Ͷ�ʱ��������Ҫ�����ߡ�������������������ʳ�����������Ӷ��������ߡ������Ͷ��Ƿ����ӵ����ʵ���Ҫ����Ҳ�в�ͬ��������϶�Ŀ������������Ͷ��в������Ľλ����˶�ѵ��ʱ�ڣ���ߵ����ʵĹ���������֯��Ԥ���˶���ƶѪ������ġ�Buskirk�ȣ�1972����ΪijЩ���ܷ������˵��˶����������ˤ�ӣ���������Ҫ������ÿ��2.5g��kg-1���ء�ʵ���о������ҹ�����˶�Ա��������Ҫ����1.8g��kg-1���أ�����2/3�����ʵ��ף�������ռ�����ܵ�12��14%����ͯ���������˶�Ա����λ���ؼ��㣬�����ʵ���Ҫ���ȳ����˸ߣ�9��11������˶�Ա����Ҫ��Ϊ3g���ҡ���֪���ҵļ��������Ӻ�ϸ�����ƻ���Yamajiͨ��ʵ��֤�������˶�ʵ�鿪ʼʱ����ϸ����Ѫ�쵰�ס�Ѫ�嵰���½����½�����ʱ��ij��̺͵�������ȡ���йأ���ÿ����ȡ������1��1.5g��kg-1���أ�Ѫ�쵰��Ѫ�嵰���ͺ��ָ���Yoshimura�������˶��ڼ���ֵ�ƶѪΪ�˶���ƶѪ�������ݶ���ʵ���Ʋ��˶���ƶѪ�Ļ����ǣ������˶�ʱ�����������������ص����ã�ʹƢ�������ͷų���Ѫ�������ƻ���ϸ�����Ӻ�ϸ���������Ѫ�쵰���������켡�쵰���µĺ�ϸ��������Ӧ�˶�����Ҫ��Ϊ��Ԥ���˶���ƶѪ��Yoshimura��������������ڼ䣬�����ʹ�����ӦΪÿ��2g��kg-1���ء�
��ʧ�ߡ�������š�������ɸı��Ӧ������£���������Ҫ������6��12%���ȣ����������ϴ�
2.6��������Ӫ��״��������
���۵�����Ӫ��״����ָ����Ҫ���������֡�
2.6.1���ϱۼ�Χ��arm muscle circumference,AMC�����ϱۼ�����arm muscle area,AMA��
�ϱۼ�Χ���ϱۼ������������嵰�״���ĽϿɿ���ָ�ꡣ�����ϱ�ΪԲͲ���ϱ۹Ǿ����ƣ������ϱ��е㴦��Χ����AC������ͷ����Ƥ��ȣ�TSF�������ɼ����ϱۼ�Χ���ϱۼ����������ʽ��
AMC��mm��=AC(mm)-3.14��TSF(mm)
AMA(mm2)=[AC(mm)-3.14��TSF(mm)]2/(4��3.14)
AMC���۱������ʱ�25.3cm(��)��23.2cm(Ů)���ձ�24.8cm���У���21.0cm(Ů)���ⶨֵ��90%��ֵΪ�������ҹ�ij��λ����1532��ͧ��Ա���У��IJ��������AMC��237mmΪ��������237mmΪȱ����AMA��4490mm2Ϊ��������4490mm2ȱ����
�ϱۼ�Χ�����㣬���۽��������������Ӫ��״�������ۣ�����K�ͼ���CT���Ľ����������ء������������������ϱ��Ƿ���εģ���ʹͬһ�˲������ϱ�Χ��Ƥ��������������ĺϼƿ�Լ��10%�����⣬���ϱ�ΪԲͲ�Ρ����Ǿ����ơ��ļ����Dz����ġ�
2.6.2��Ѫ�嵰��
�͵����ſ�����Ѫ�����ϳɽ��͡���Ѫ�����׳�ȥ����plasmaphresis��ʹ���ﶪʧ50%��Ѫ�����ף������Ըߵ����ſ���һ����Ѹ�ٻָ�����ʧ��1/3�����ڶ�������ȫ�ָ�������ˮƽ�����Ѫ���ܵ��Ͱ��׳��ڵ�������ֵ������˵���嵰�ײ��㡣
Ѫ�嵰���а��ף�albumin,Alb����ǰ���ף�prealbumin,PreAlb�����������ף�transferrin,TFN�����ӻƴ���ϵ��ף�retinol binding protein,RBP����Ҫ���ڸ���ϳɡ��⼸��Ѫ�嵰��Ũ�Ƚ��ͣ�������Ϊ����������ȱ���������ϳɼ��͵�Ե�ʡ�
��1�����װ�����Ⱥ�����ʱ���õ�ָ�ꡣ��Ⱥ���鷢��ƽ��Ѫ�����ˮƽ�ͣ���������ʳ���������������йء�
Alb���۱�����35g��L-1������28��34g��L-1���ȱ����21��27g��L-1�ж�ȱ������21g��L-1����ȱ���������ʵ���Ũ�ȵ���28g��L-1ʱ�������ˮ�ס�
���ײⶨ��Ʒ�ײɼ����������ס�����������4��5g��L-1������������ڣ�20d����������ȱ��ʱ���ײ����
��2���������������������������ĵ��ס��Ͱ��ױȽϣ�������������С����������ڣ�8��10�գ��϶̣����ܼ�ʱ�ط�ӳ����������ı仯���ڸߵ���������ʱ��Ѫ����Ũ�������죬���ж�����Ч��������ָ�ꡣ
TFN���۱���2500��3000mg��L-1������1500��2000 mg��L-1���ȱ����1000��1500mg��L-1�ж�ȱ������1000 mg��L-1����ȱ�����÷������߷��ⶨ����
��������Ũ����������Ӱ�졣�������ʺ�������ȡ������ʱ����Ѫ��Ũ�ȳ��ִ��������ߣ�������ʱӦע�⡣
��3��ǰ����ǰ������Ҫ�����������״���ء���������С�����������1.9�졣
PreAlb���۱���157��296 mg��L-1Ϊ������100��150mg��L-1���ȱ����50��100 mg��L-�ж�ȱ������50 mg��L-1����ȱ����
���κμ���ϳɵ����ʵ�����£��紴�ˡ����Ը�Ⱦ��Ѫ��ǰ����Ѹ���½�������Ӳ��������ж��Ƿ��е�����Ӫ�������������ء�
��4���ӻƴ���ϵ����ӻƴ���ϵ���������ά����A�������⵰�ס�����С���˹����������л�����������10h�������۵�����Ӫ���������Ա仯������ָ�ꡣ
RBP���۱���2��76 mg��L-1Ϊ��������ָ��߶����У������ں�С��Ӧ������£�Ҳ�б仯������ٴ�����Ӧ�á������в���ʱ��Ѫ��RBPŨ�����ߡ�
���⣬Ѫ���ܵ��ס���Ҳ��������ָ�ꡣ�ҹ�����������Ѫ���ܵ�������ֵ��65��80g��L-1������/�ı���1.5��2.5:1����������ָ�������Բ�������ף����и�Ⱦ�ͼ����没ʱ�����ߡ�
Ӧ�ÿ�����Ѫ�嵰��Ũ�Ȳ����뵰������ȡ�ͺϳ��йأ�Ҳ�ֽܷ⡢Ѫ���������С�������ϸ����Һ���ӵ����ص�Ӱ�졣��ˣ�������ʱ�����ۺϷ�����������ڼ��½��ۡ�
2.6.3��Ѫ�就�����ֵ��serum amino acid ratios,SAAR��
�ڵ�����Ӫ������ʱ������������Ӧ�Դ�л�Ľ����Ѫ�����백�����ģʽ�����仯��������Ӫ�������Ķ�ͯ���ո�Ѫ�����ᡢ��������ȱ��谱������Ұ��ᡢ������ȷDZ��谱������٣��������DZ��谱�������������ߡ�
SAAR=���ʰ���+˿����+������+ţ���ᣩ/����������+������+�Ӱ���+�����ᣩ
���۱���SAAR��2Ϊ��������3������Ӫ��������
��ָ������������ӣ����������߱����������������������ʲ���������£��������壬����������á�
2.6.4����������urinary creatinine��
���������Լ�������ἡ�ᣬ�������ߴ��ڹǸ켡���ο�2.2.3�ڣ�����ˣ��ⶨ24h�������Ϊ������֯Ӫ��״�����۵�ָ�ꡣ��Ⱥ�������������֯�������������зdz���������ء������ڼ�������ʳ������ںϳ��������֣����������ۏC����Լ��0.3��0.5%�����ڲⶨ�����ϣ�24h��������ȷ�ռ�����ʹ����ʱ����15min֮�Ҳ��ʹ�ⶨ�������1%��
2.6.5��������/����ָ����urinaryceratinine/height index,CHI��
������/����ָ����24h��������A����ͬ�Ա�ͬ���ߵij�����24hԤ�ڵ���������B���ı�ֵ��
CHI=A/B��100
A=������24h������
B=��������ͬ�����е������������ص���Ԥ�ڵ�24h����ֵ[����ü�����ϵ��23mg��kg-1(��)��18 mg��mg-1(Ů)���ؼ���]��
CHI���۱���60��80%�ж�ȱ������60%����ȱ����
����ָ�������嵰��Ӫ��״��������һЩ���⣺��24h������ȷ�ռ������������ز����ã���Щ��������ʱ�����ز������������أ��۱�û�������𣬶�ʵ�������������������������͡�����ٴ�Ӧ�ü�ֵ����
2.6.6����������������̬���
ͷ������ʱ��ë����ϸ���и߶Ⱥϳɵ����ʵ���������ֳ�졣���ͷ������������̬������Ϊ�����жϵ�����Ӫ��������ָ�ꡣ��Zain���˵ĵ��飬335����ͯ������DNA�Ϳ����Ե������浰����Ӫ�������ķ�չ���н����Եļ��١�Bradfield(1970)�۲�8���������ӵ����ʵ���ȡ��ͷ�������Ĺ�ϵ������ʳ������15���������������ή������Ѫ�嵰����δ�����쳣��Smelser�����ð�ë���Ʋⶨ�η�����Ĺ������Է�ӳ������Ӫ��״������ë�����Ⱥ����ء��ϱۼ�Χ��Ѫ�������������ء�
����ָ�꣬������Ȼ�ܶ࣬�����в���֮����ʵ��Ӧ�û�������ʳʷ���ٴ��۲�����ۺ����ۡ�
2.7��ʳ�ﵰ����
2.7.1��ʳ�ﵪ�Ĵ�����ʽ
ʳ����Դ�ڶ���ֲ����е��Ĵ�����ʽ�кܴ���������ֵĵ��Ե�������ʽ���ڣ������������백��������Լ����ᡢ��֬�������ᡢ�켡�ģ�������ǵ��������ḻ��Լռ�ܵ�����10��30%���鵪Լ20%���ڷǵ�����ֲ����ʳ�ﺬ��������ijɷֲ�����������༸��95%�ĵ������ڵ����ʣ��������������������ܲ��ȣ�����������50%�����������ĺ����백�������ʽ���ڣ��ر������������Ȱ��������Ŷ����ᡣ���⣬ֲ����֯�к��в��ٷǵ��װ����ᣬ��Щ��������Щ�������ڴ�л��������ԭ����������ų��������������ж��ģ��������е��������ᡢ�϶��еĦ���������ᡣ�µĵ�������Դ�絥ϸ�������˵��ܸߣ������ɴﵰ����������50%�����ں��������������մ�л����Ϊ���ᣬ����ʳ�ÿ�����Ѫ������Ũ�����ߣ����γ����ʯ��ʹ��֢����ˣ���ϸ�����ײ���ȥ���ᴦ����������Ϊ����ʳ�����Դ��
ʳ���е����ʵĺ���һ����ÿ��϶��������вⶨ��Ȼ����ɵ����ʵ���������ֲ����ʳ�ﵰ�ĺ�����ԼΪ15.7��19%��ƽ��16%������õĵ�ֵ����6.25�������ʻ���ϵ���������ø�ʳ��Ĵֵ��ĺ�������Ҫ�Ƚ�ȷ�ؼ���ʱ���ɲ��ò�ͬ�Ļ���ϵ������2-7����
��2-7 ������ɵ����ʵĻ���ϵ��
| ����ϵ�� | |
| ��ۣ��л�ͳ����ʣ� | 5.70 |
| ȫ�� | 5.83 |
| ���� | 5.95 |
| ���� | 5.46 |
| �ƶ� | 5.71 |
| ֥�� | 5.30 |
| ���� | 6.38 |
���ּ��㷽���ǰ�ʳ���ܵ�ȫ���Ե����ʵ���ʽ���ڶ�����ģ��Ժ��ǵ����ߵ�ʳ����˵�������ʵļ����ֵ���ɹ��ߡ�����ö�������õĵ����ʣ���֮Ϊ�ֵ��ס�
2.7.2������ʳ��ĵ����ʺ���
ʳ��ɷֱ���ʳ��ĵ����ʺ�������ÿ100gʳ���е�����ʾ�ġ������û�б����ʳ��ĵ����ʺ����ܵĹ�ϵ����Ϊ�����������Ҫ������ʳ�����ȡ������ˣ�ʳ����Ϊ��������Դ�ļ�ֵҲ�������䱾������ֵ�����ҹ��������Ӵ����������Ͷ�ʱ��ÿ����ʳ���ܹ�����Ϊ10920kJ�������ʹ�����Ϊ80g����ʳ�ﵰ���ṩ������Լռ�����ܵ�11%�����˵�ʳ����е������ṩ������ռ�����ܵ�10��15%����ˣ���ʳ���е������õ����ʵ�����ռʳ�������ܵİٷ�����ʾ����2-8�������Դ����жϸ�ʳ����Ϊ��������Դ�ļ�ֵ���ӱ�2-8�����������ƶ����㡢�����ⶼ�Ǻܺõ�ʳ�ﵰ����Դ�����ѡ�������Ϊ��ʳΨһ��ʳ����Դ���䵰������Ȼ�����������嵰���ʵ���Ҫ����
��2-8 ����ʳ��ĵ���������������ʳ�������ܵıȣ�%��
| ������ | ||
| g��100g-1ʳ��* | kJ��100kJ-1ʳ��** | |
| ƻ�� | 0.3 | 2.8 |
| ���ף��ϰ��� | 6.7 | 7.8 |
| ���� | 18.1 | 52.1 |
| С��ۣ���ǿ�ۣ� | 9.4 | 10.7 |
| ���� | 2.3 | 11.9 |
| ������ | 26.2 | 19.2 |
| ������ | 16.7 | 20.2 |
| ���� | 14.7 | 34.6 |
| �ƶ� | 36.3 | 35. |
| ���������� | 7.4 | 41.1 |
| ţ�� | 20.1 | 46. |
*ժ���й�ҽѧ��ѧԺ�����о���:ʳ��ɷֱ�
**��ʳ��ɷֱ�����ó�
2.7.3����ʳ�����ʵ���������
��ʳ�ĵ����ʵ�Ӫ����ֵ�ںܴ�̶��ϣ�ȡ����Ϊ����ϳɺ��������������ṩ���谱���������ģʽ���������������������ķ��������Դ˸�����Ϊ�����ġ����۵ķ����������֣����κ�һ�ַ�������һ��������Ϊ����ָ�꣬�������һ���ľ����ԣ�����ʾ��Ӫ����ֵҲ����Եģ���ˣ���������һ��ʳ�����ʳ�ﵰ��ʱ��Ӧ�ø��ݲ�ͬ�ķ����ۺϿ��ǡ������������ֳ��õ����۷�����
��1�������������ʣ�digestibility,D��ʳ��ĵ�������������ָʳ�ﵰ��������øˮ������յij̶ȣ������յ������ܵ����ı�ֵ��ʾ��
D=����N/����N��100
ʳ�ﵰ������ʵ�����ʣ�ture digestibility,TD�����ý�ʳʵ���ã�
TD=[����N-����N-���лN��]/����N��100
�൪��ȫ��δ������ʳ�ﵪ��������һ�����������䳦ճĤϸ��������ø�ͳ�������ⲿ�ֵ���Ϊ���л����������������ʳ������ʱ����÷൪��֪������ԼΪ0.9��1.2g��24h-1��������л�����Բ��ƣ���Ϊ����������(apparent digestibility,AD):
AD=(����N-��N)/����N��100
���������ʱ���ʵ�����ʵͣ��Ե�����Ӫ����ֵ�Ĺ���ƫ�ͣ�����нϴ�İ�ȫϵ�������⣬���ڱ��������ʵIJⶨ������Ϊ��㣬��һ�����á�
��һ����������ӹ���ʳ�ﵰ��������Ϊ������97��98%������92��94��������98%������82%������74%��ֲ����ʳ�ﵰ����������ά��Χ���ȶ�����ʳ�ﵰ��������Ҫ�ͣ�����ά�ؾ��ӹ������ƻ����ȥ��ֲ�ﵰ�������ʿ�����ߡ������������Ϊ60%���ӹ��ɶ�������ߵ�90%��
��2�������ʵ������ֵ��biological value,BV�������ʵ������ֵ��Ϊά�ֺ�/�������������ڱ����������յ��ı�ֵ��
BV=[����N-����N-���лN��-����N-����ԴN��]/[����N-����N-���лN��]��100
�����������ֵ�ܺܶ����ص�Ӱ�졣�Բ�ͬʳ�ﵰ�������ֵ���бȽ�ʱ��ʵ������Ӧ��һ�£�����ʹͬһ��ʳ��Ҳ��ó���һ�µĽ�����缦����������ռ������8%ʱ�������ֵΪ91��ռ16%ʱΪ62��һ������£�ʵ�鶯�����ó�����Ĵ��������е����ʺ���ռ10%������ʳ�ﵰ�������ֵ����2-9��
��2-9 ����ʳ�ﵰ�������ֵ
| �����ֵ | �����ֵ | �����ֵ | |||
| ���� | 77 | ���� | 67 | ȫ���� | 94 |
| С�� | 67 | �� | 64 | ţ�� | 76 |
| ��� | 52 | �϶� | 58 | ���� | 74 |
| ���� | 72 | ���� | 59 | Ϻ | 77 |
| ���� | 60 | �ײ� | 76 | ţ�� | 85 |
��3�������ʾ������ʣ�net proteinutilization,NPU�������������ֵû�п���������������δ���ն���ʧ�ĵ�������Miller�Ƚ��齫�����ֵ���������ʣ���֮Ϊ�����ʾ������ʣ�
NPU=BV��D=����N/����N
����ĵ����ʾ�������Ҳ�����嵪�����вⶨ����ͬ�Ѷ������ֱ����Ժ�ά��ˮƽ�����ʵ�ʵ�����ϣ�A�飩���������ϣ�B�飩��10d����¼����ÿ����ʳ����ʵ������ʱ�ⶨ���鶯��ʬ���ܵ��������Ϻ������������м��㣺
NPU=��BF-BK+IK��/IF
ʽ��: IF=A��N������
BF=A��ʬ����N��
BK=B��ʬ����N��
IK=B��N������
Ҳ�ɲⶨʵ�鶯��ʬ��ĸ��غͺ�ˮ����������֪������ʬ��N/H2O��ƽ����ֵ����ʬ�庬�������������Ϊ��㡣
��4��������Ч���ȣ�protein efficiencyratio,PER��������Ч���������뵥λ���������ʵ�������������
PER=�������ӣ�g��/���뵰���ʣ�g��
ͨ�������Զ������Ϊʵ�����Osborne��֤��PER�������е����ʵ�ˮƽ���ı䣬������������˵ĵ����ʵ�ˮƽ�Ͻ���ʵ�顣ϰ�����ú�10%�����ʵ����ϣ�AOAC����ı��������ú�9.09%�����ʵ������������
�˲ⶨ����ȱ����û�а�ά������ĵ����ʿ������ڣ�������ý�������ɱ���������PERΪ2ʱ��������������PERΪ1ʱ����������ͬʵ���õ�PER���ظ����������ѣ�Ϊ�˼���ʵ���Ҽ�ı��죬�����ҵ��ף��ο����ף���PERֵ��
У����PER=PER����2.5/�ҵ���ʵ��PER��
��5����Ե����ʼ�ֵ��realative proteinvalue,RPV����Ե����ʼ�ֵ�Ƕ�����ʳ���Ե��ļ���-��������б�ʣ�A������ʳ�ο����ļ���-��������б�ʣ�B���ȣ�
RPV=A/B��100
�Ժ�3��4�ֲ�ͬ����������ʳ�ﵰ��ι�������������������������Y�������Ե��Ľ�ʳ������X�����ƻع鷽�̣�����б�ʣ�A����ͬʱ�ú���ͬ����������ף��ο����ף�ι�����ͬ���ü���-�����ع鷽�̼�б�ʣ�B��������ǰһ�ع鷽��ΪY1=2.35X1-0.36����һ�ع鷽��ΪY2=4.12X2-0.28��������Ե�����Ե����ʼ�ֵ�ɼ������£�
RPV=2.35/4.12��100=57
�����Ե��ײ�õĻع鷽�̣�б��Խ����������Խ�ߡ�
��6�����������֣�amino acid score����ѧ���֣�chemicalscore�� 1946��Block��ָ���ںϳɵ����ʵij��������ɵ�����������İ����ᣨAA������ͬʱ���ڣ�ȱ�������κ�һ�־ͻ�Ӱ��ϳɣ������ʳ�ﵰ�װ������������۵����ʡ���������ⶨij������ʳ�ﵰ����ʳ�ﵰ����ÿһ�ֱ��谱����ĺ�������ο������бȽϣ���ÿ�ְ�������ο����װ�����ı�ֵ��ʾ����ֵ��͵����ְ����ᣬ��Ϊ��һ���ư����ᣬ����ͱ�ֵ������ʳ�ﵰ�İ��������ֻ�ѧ���֡����������ֿɼ������£�
����������=ÿ�����Ե���ij��AA������mg��/ÿ�˲ο����ĸ���AA������mg����100
�� ����������=���Ե���ÿ��N��ijAA����(mg)/�ο�����ÿ��N�ĸ���AA������mg����100
ʳ�ﵰ���а�����ĺ������ñȽϵIJο����Ĺ�ϵ��ʾ����ÿ�˵��Ĺ�ϵ��ʾ��Block��ԭ���ѵ�����Ϊ�����Ե��ױȽϵIJο����ף����������ֶ����FAO/WHO��1973������ѧ��ǰ��ͯ�����Ҫ���ƶ������백������Ҫ��ģʽ����2-10�������Ե����бȽϡ�
��������ģʽ��õİ��������������ڶ�ͯ��ʳ�ﵰ�������ʵ����ۣ����Գ���������˸õ����ʵ���������Ϊ�����˶�ÿ�˵����ʰ��������Ҫ���ϵ͡�
��2-10 ����İ�������Ҫ��ģʽ
| ������mg��g-1�����ʣ� | |
| �������� | 40 |
| ������ | 70 |
| ������ | 55 |
| ������+�װ��� | 35 |
| ��������+�Ұ��� | 60 |
| �հ��� | 40 |
| ɫ���� | 10 |
| �Ӱ��� | 50 |
��2-11 ����ʳ�ﵰ�İ���������
| ��������Դ | �����Ậ����mg��g-1�����ʣ� | ���������֣����ư����ᣩ | |||
| ������ | ������ | �հ��� | ɫ���� | ||
| ����ģʽ | 55 | 35 | 40 | 10 | 100 |
| ���� | 24 | 38 | 30 | 11 | 44�������ᣩ |
| �� | 72 | 24 | 42 | 14 | 68�������ᣩ |
| �̷� | 80 | 29 | 37 | 13 | 83�������ᣩ |
| �ȡ������̷ۻ�ϣ�67��22��11�� | 51 | 32 | 35 | 12 | 88���հ��ᣩ |
*����Pellett,Pl etal(1978):��Nutritional Evaluation of Protein Foods������
2.7.4�������ʵĻ�������
����ʳ���ʳ�����ڱ��谱�����������������ಹ�䣬���ܸ��ӽ�������Ҫ���ı�ֵ��ʹ�����ֵ�õ���Ӧ����ߣ����������Ϊ�����ʵĻ������á���С��С�ס�ţ�⡢��������ʳ��ʱ���䵰���������ֵ�ֱ�Ϊ67��57��69��64������ʳ�������ֵ�ɸߴ�89�������Ͱ�����Ӫ���о�����INCAP���Ƴ�һ��ֲ����ʳ���Ϊ��Incaparina�������к�����29%������29%�����ѷ�38%��ơ�ƽ�ĸ3%��̼���1%��ά����A�����ֻ��ʳ����Ӫ�����������͵�����ʳ�����ò���ʳ��䵰���ʵ������ֵ���Դ���ţ�鵰�ס�
�����ư����Ჹ�䵽��Ӧ��ʳ���У����������Ჹ����൰�ף��õ����ᡢ��������հ��Ჹ�仨���ۣ�ͬ�������������á��������������������0.2%����۵��������ֵ����47��ߵ�71��ѧ���ͯʳ������������ǿ��ʳƷһ������ߡ����غ͵ֿ����Ⱦ��϶�������������ߡ�
��Ϊ��ɵ����ʰ��������ͬʱ���ڲ��ܺϳɵ����ʣ����һ����ڰ�����Ĵ��������٣������ʳ�в�ͬ�����ʱ�����ͬһ����������������á���ÿ3h������һ�ֱ��谱������������������ò��ѣ�������������
2.7.5���ӹ���ʳ�ﵰ��Ӫ����ֵ��Ӱ��
ʳ��ӹ��ķ����м��ȡ��䶳�����衢��ѹ������ȣ������Լ��ȶԵ����ʵ�Ӱ��������ʾ������ȴ��������ͣ�Configuration���ı䣬���е��������ɥʧ�����ֱ仯��Ϊ���ԡ��絰���������̡���������������Ӳ���DZ��������ֵ����ʵ��������ܲ�һ��������60��80�濪ʼ���ԡ����Բ�ͬ�ڱ��ʣ������ʵ�һ���ṹδ�䡣
����ͷ�ֹʳ�︯����������100��200��ļ��ȷ����������¶��º�û���Ǵ���ʱ�������ʷ������ԣ�ά�ֵ����ʿռ乹��Conformation���Ĵμ����������ѣ��ƻ����ļ�ԭ�еĿռ����С�ԭ���ڷ����ڲ���һЩ�Ǽ��Ի��ű�¶�����ӱ��棬ʹ�����ʵ��ܽ�Ƚ��ͣ��������̡�ͬʱ���ַ�Ӧ������-NH2��-COOH��-OH��-SH�ͷų�����ʹ����������ø�⣬Ҳ�������������ʳ���а��������ʧ����
ijЩʳ���к����谭ø���õ����Ƽ�������еĿ��ȵ���ø��Ѫ�������أ������е���ճ�������Ⱥ�����Զ�ʧȥ���ԡ�����˶�ø���������ã��Ӷ������ʳ���Ӫ����ֵ��
��ʳƷ���������⣬�����о�ԭ�Ե����ࡣ�����ʹ��ȼ��ȣ��������л�ԭ�Ǵ��ڵ������£��ɲ�����ø�������£�Maillard����Ӧ��ʳ�����غ�ɫ�����а�������Ҫ����������ƻ��������˵����ʵ������ֵ��ͬʱ�����ʵ�ø��Ҳ�½���ʹʳ�ﲻ�������������·�Ӧ�Ĺ�����Ϊ���ӣ���ʹ�ڽϵ��¶���Ҳ�ܽ��У�ֻ�Ƿ�Ӧ�����൱������
�����
1��Munro HN andCrim MC:In��Modern Nutrition in Health and Disease��,Goodhart RS Ed.pp51��98,1980
2.���ɷ���Ӫ��ѧ��3��4����185��1981
3����ѧ��ȣ�������ȫ��Ӫ��ѧ����������ժҪ��࣬1ҳ��1981
4��������ȣ�ʵ���˶�ҽѧ�����棩���������������磬152ҳ��1982��
5��Grant TP etal:Surg Clin N Amer 61(3):437��464,1981
6.SmelserDN:AmJ Clin Nutr35(2):342,1982
-
���ٴ�Ӫ��ѧ�� �е�����½ڣ�
��һƪ��Ӫ������
����1������
����2�������ʣ���ǰҳ��
����3��֬��
����4��̼ˮ����
����5������
����6�����Σ�����Ԫ�أ�
����7����Ԫ��
����8��������Ӫ��
�ڶ�ƪ��Ӫ��ȱ����
������