第二节 白细胞分化抗原的应用
CD抗原及其相应的单克隆抗体在基础和临床免疫学研究中已得到广泛的应用。在基础免疫学研究中,CD主要应用于:(1)CD抗原的基因克隆,新CD抗原及新配体的发现;(2)CD抗原结构与功能关系;(3)细胞激活途径和膜信号的传导;(4)细胞分化过程中的调控;(5)细胞亚群的功能。在临床免疫学研究中,CD单克隆抗体可用于:(1)机体免疫功能的检测;(2)白血病、淋巴瘤免疫分型;(3)免疫毒素用于肿瘤治疗、骨髓移植以及移植排斥反应的防治;(4)体内免疫调节治疗。有关与免疫功能相关的CD分子归纳于表1-4。有关与T细胞表面分子、B细胞表面分子以及NK细胞的表面标记参见第七章。与CD有关的Ig超家族、粘附分子、补体受体、细胞因子受体等分别在本书的有关章节中加以介绍。
表1-4 与免疫功能有关的CD
| 免疫功能 | CD |
| 细胞受体 TCR CD3、CD4、CD8 mIg mb-1/Igα(CD79a)、B29/Igβ/(CD79b) CR CR1(CD35)、CR2(CD21)、CD3(CD11b/CD18)、CR4(CD11c/CD18)、C5aR(CD88) FcR FcγRI(CD64)、FcγRⅡ(CD32)、FcγRⅢ(CD16)、FcεRⅡ(CD23)FcαR(CD89) 细胞因子受体 IL-2Rα(CD25)、M-CSFR(CD115)、GM-CSFR(CDw116)、SCFR(CD117)、 IFN-γR(CDw119)、TNF-αR(CD120)、IL-1R(CDw121)、Il -2Rβ(CD122)、 IL-4R(CDw124)、IL-6R(CD126)、IL-7R(CDw127)、IL-8R(CDw128)、 gp130(CDw130) 细胞间、细胞基质相互识别 白细胞粘附分子-内皮细胞粘附分子: LFA-1(CD11a/CD18)-ICAM-1(CD54)、ICAM-2(CD102) Mac-1(CD11b/CD18)-ICAM-1(CD54) VLA-4(CD49d/CD29)-VACM-1(CD106) L-selectin(CD62L)-E-selectin(CD62E)、P-selectin(CD62P) CD15-E-selectin(CD62E)、P-selectin(CD62P) 淋巴细胞归位受体-血管内皮细胞地址素: L-selectin(CD62L)-PNAd CLA-E-selectin(CD62E) LFA-1(CD11a/CD18)-ICAM-1(CD54)、ICAM-2(CD102) VLA-4(CD49d/CD29)-VCAM-1(CD106) CD44-MAd LPAM-2(CD49d/β7)-MAd、VCAM-1(CD106) 白细胞粘附分子-细胞外基质: VLA-1(CD49a/CD29)-CA、LM VLA-2(CD49b/CD29)-CA、LM VLA-3(CD49c/CD29)-FN、LM、CA VLA-4(CD49d/CD29)-FN VLA-5(CD49e/CD29)-FN VLA-6(CD49f/CD29)-LM α7β1(-/CD29)-LM VNR-β1(CD51/CD29)-VN、FN Mac-1(CD11b/CD18)-FB P150,95(CD11c/CD18)-FB GPⅡbⅢa(CD41/CD61)-FB、FN、vWF、TSP VNR(CD51/CD61)-VN、FB、vWF、FN、CA、TSP α6β4(CD49f/CD104)-LM VNR-β5(CD51/-)-VN CD51/ β6-FN CD49d/β7-FN GPIb-α/IX(CD42b/CD42a)-vWF 免疫细胞间相互识别: CD22-CD45RO CD2-LFA-3(CD58) CD4-MHCⅡ类分子 CD5-CD72 CD 8-MHCⅠ类分子 LFA-1(CD11a/CD18)-ICAM-1(CD54)、ICAM-2(CD102) CD28-B7/BB1(CD80) CD27-CD70 参与白细胞激活 T细胞:CD2(T细胞旁路激活途径)、CD3(信号转导)、CD4、CD5、CD8、CD28、 CD43、CD44、VLA-4(CD49d/CD29)、CDw90 B细胞:CD19(抑制G0→C1,抑制Ig分泌)、CD20(抑制细胞周期)、CD21(活化B细 胞)、CD22(Ca2+升高,促进G0→G1)、CD23(B细胞分化)、CD40(B细胞生 长)、CD72、CD73(G0→G1)、CD80(B细胞活化) 髓样细胞:CD14(髓样细胞氧化爆发?)、CDw65(中性粒细胞活化) NK:CD16(NK活化)、CD2、CD3 非谱系:CD69(活化诱导分子,AIM) 与细胞膜表面酶有关 CD10(中性肽链内切酶)、CD13(氨肽酶)、CD26(二肽酰肽酶Ⅳ)、 CD45(酪氨酸磷酸酯酶)、 CD73(5`核苷酸外切酶)、CDw75(具有酶活性) 与病毒受体有关 CD21(EB病毒R)、CD4(HIVR)、CD54(鼻病毒R)、CD46(麻疹病毒受体) | |
一、与T细胞识别、粘附、活化有关的CD分子
T细胞是一类重要的免疫活性细胞,除直接介导细胞免疫功能外,对机体免疫应答的调节起关键作用。T淋巴细胞本身的识别活化及效应功能的发挥,不仅与外来抗原、丝裂原和多种细胞因子密切相关,而且有赖于T细胞相互之间、T细胞与抗原提呈细胞(APC)之间以及T细胞与靶细胞之间的直接接触。T淋巴细胞识别抗原的受体是T细胞受体(t cell receptor,TCR)与CD3所组成的复合物(TCR/CD3)。在识别过程中还有赖于抗原非特异性的其它细胞表面分子的辅助,这些辅助分子(accessory molecules)主要包括CD4、CD8,MHC Ⅰ类分子、Ⅱ类分子,LFA-1(CD11a/CD18)、CD49d、e、f/CD29(VLA-4、VLA-5、VLA-6)、CD28、CD44、CD45、ICAM-1(CD54),LFA-2(CD2)和LFA-3(CD58)等。
有关MHCⅠ类、Ⅱ类分子的结构和功能在第六章“MHC及其临床应用”中讨论。VLA-4、VLA-5、LFA-6、LFA-1、ICAM-1、CD44见第二章“粘附分子”。有关CD45在第八章“免疫球蛋白超家族”中阐述。

图1-1 参与T细胞对靶细胞识别的分子(模式图)
T细胞表面的辅助分子有以下特点:
(1)存在于T细胞上的辅助分子可特异地与存在于APC或靶细胞上的某些分子(配体)相结合,如LFA-1和CD2可分别与ICAM-1和LFA-3结合。
(2)辅助分子本身不具有多态性,在一个物种所有个体的所有T细胞的某一种辅助分子的结构基本上是相同的。
(3)辅助分子可加强T细胞与APC或靶细胞结合的程度。
(4)许多辅助分子具有转导信号的功能,如CD2、CD4和CD8等分子。
(5)有些辅助分子如CD2、CD4、CD8、CD28、Thy-1等其编码的基因属于Ig基因超家族;有些辅助分子如LFA-1、VLA-4、VLA-5和VLA-6等编码的基因属于integrin 基因超家族。
(6)T细胞膜表面辅助分子作为膜表面重要的标记已被应用于临床的诊断和治疗。
(7)细胞因子可调节辅助分子的表达,从而改变细胞间粘附的能力,这是细胞因子免疫调节作用的一个重要方面。
(一)T细胞受体
T细胞受体(T cell receptor,TCR或Ti)是T淋巴细胞表面识别外来抗原与自身MHc Ⅰ类抗原(或Ⅱ类抗原)复合物的受体,在同种异体移植中TCR也识别单独的非已的MHC抗原。目前已经证实,TCR在细胞表面与CD3密切结合在一起组成TCR/CD3复合物,TCR识别抗原后刺激信号是通过CD3分子传递的。
1.T细胞受体的类型和结构 TCR中的多肽链是异质性的。根据抗原结构和编码基因不同,已发现有α、β、γ和δ四种多肽链。关于TCR多肽链的结构大多是从分析TCR多肽链cDNA或基因组克隆(genomicclones)而来,编码TCR多肽链的基因属于免疫球蛋白基因超家族成员。成熟TCR肽链分子量在40~60kDa之间。根据TCR中异源双体的组成的不同,TCR可分为以TCRαβ和TCRγδ两种类型。
(1)TCRαβ:CD阳性TCRαβT细胞可识别非已MHCⅡ类抗原(同种异体抗原)或自身MHCⅡ类抗原与加工后抗原的复合物CD8阳性TCr αβT细胞则可识非已MHCⅠ类抗原或自身MHC Ⅰ类抗原与加工后抗原的复合物TCRα链分子量40~50kDa的酸性糖蛋白,β链40~50kDa不带电或碱性糖蛋白。α和β链各由一个可变区(V区)和一个恒定区(C区)组成,与Ig的V区和C区大小相似,属于免疫球蛋白超家族成员。TCRα、β链的V区约含102到109个氨基酸,在V区部分由两个半胱氨酸形成链内二硫键,组成约含50~60氨基酸残基的环肽,这与IgV区结构和功能相似,是特异性识别外来抗原的结构域。TCRα、β链的C区约含138到179个氨基酸,每个C区形成由链内二硫键连接的环肽。α、β链在连接肽(connectingpeptide)形成链间二硫键。穿膜区约由20~24氨基酸组成,α链穿膜区含有带正电的1个赖氨酸和1个精氨酸残基,β链穿膜区含有1个带正电的赖氨酸残基,这些带正电的氨基酸与CD3γ、δ和ε链穿膜区带负电的谷氨酸和/或天冬氨酸形成盐桥,稳定TCR/CD3复合物结构,并与CD3传递信息有关。α、β链胞浆部分只有5~12氨基酸长的尾部(图1-2)。
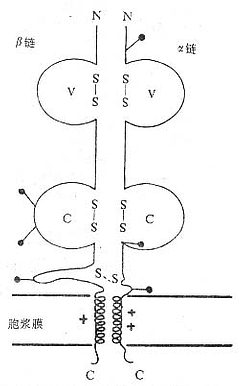
图1-2 TCRαβ异源双体模
(2)TCR γδ:TCRγ和δ链各包括一个Ig样的V区和C区、连接肽、疏水的穿膜区以及一个短的胞浆区尾部,在连接肽区可形成链间的二硫键。γ和δ链的穿膜区各含有1个带正电的赖氨酸,此外δ链还有1个带正电的精氨酸,这些带正电的氨基酸与CD3γ、δ和ε链穿膜区带负电的天冬氨酸或谷氨酸形成盐桥。在氨基酸水平上分析,TCRγ链与β链同源性较高,而TCRδ链与α链同源性较高。在人类TCRγδ有二硫键相连和非共价相连两种形式,而在小鼠只发现二硫键相连的TCRγδ形式。人γ链分子量为36~55kKa,δ链为40~60kDa,γ、δ链的分子量大小取决于多肽骨架的长度和糖基化的程度。
有关TCRα、β、γ、δ链基因的结构和重排见第三章“免疫球蛋白超家族”
2.两种类型TCR T细胞的比较 TCRαβ与TCRγδ不仅组成受体多肽链的结构不同,而且具有这两种类型受体T细胞的分布、表型、发育以及功能也有差别(表1-5)。
表1-5 TCRαβ与TCRγδ细胞特性的比较
| 特性 | TCRαβ | TCRγδ |
| |
| 分 布 | PBL | 60~70% | 0.5~15% |
|
| 其它部分 | 小鼠树突状表皮细胞(DEC)、小鼠粘膜上皮内淋巴细胞(IEL) |
| ||
| 表 型 | CD4+CD8- | 60~65% | <1% |
|
| CD4-CD8+ | 35% | 20~50% |
| |
| CD4-CD8- | <1% | 50~80% |
| |
| CD2 | 100% | 100% |
| |
| CD5 | >95% | -或+(弱) |
| |
| 发 育 | 胸腺后期 | 胸腺早期(早于TCRαβ) |
| |
| 功 能 | (1)识别MHC与加工处理 多肽的复合物 | (1)识别外来抗原的MHC限制尚有争论,识别破伤风类毒素可能受MHCⅡ类抗原限制;小鼠TCRγδ对合成肽反应受Qa分子限制。 |
| |
| (2)识别非已MHC及MHCⅠ类抗原相关分子(TLa、CD1) | ||||
| (2)识别非已MHC | ||||
| (3)产生些细胞因子(IL-2、IL-4、IL-5、GM-CSF、IFN-γ) |
| |||
| (3)产生多种淋巴因子 | (4)某些TCRγδ细胞可杀伤靶细胞 |
| ||
| (5)调节TCRαβ的发育。 | ||||
| (4)杀伤病毒感染等靶 细胞 | ||||
| (6)上皮屏障(针对肠毒素、分枝杆菌热休克蛋白等) |
| |||
注:树突状表皮细胞dendriticepidermal cell, DEC
上皮内淋巴细胞intraepithelial lymphocyte IEL
在正常外周血中,CD4-CD8+、CD4+CD8-、CD4+CD8+和CD4-CD8-四种表型不同的T细胞分别占T细胞总数的25%、70%、1%和4%左右,其中前三种表型TCR类型主要为TCRαβ,而CD4-CD8-T细胞主要为TCRγδ。以下疾病可见外周血或局部TCRγδ细胞数量或比率升高:(1)重症联合免疫缺陷、常见可变型免疫缺陷、Wiskott-Aldrich综合征、Di-George综合征、白血病患者骨髓移植等病人外周血中TCRγδ细胞百分率增加;(2)少数急性T细胞白血病、T细胞恶性淋巴瘤患者为TCRγδT细胞发生恶性变;(3)慢性淋巴细胞性白血病、大颗粒淋巴细胞(LGL)白血病病人PBMC中TCRγδT细胞百分率增加;(4)肾移植患者排异反应晚期外周血中TCRγδ细胞增加;(5)类风湿性关节炎患者关节腔滑液中TCRγδ阳性细胞比率要高于外周血中TCRγδ细胞比例,推测TCRγδ可能参与局部炎症的发生;(6)经结核杆菌免疫后的局部淋巴引流液中TCRγδT细胞比例增加,麻风结节病灶中有很高比例的TCRγδT细胞,提示TCRγδT细胞对分枝杆菌所致的感染免疫中起重要作用;(7)HIV、EBV感染时外周血TCRγδT细胞比例增加。
[超搞原]有的抗原不经过APC处理和递呈可直接激活CD4阳性T细胞称为超抗原(superantigen,SAg),具有类似致分裂原的作用。SAg对T细胞的激活采取一种独特的方式,即分子一端和TCRβ链上V基因产物结合,别一端和APC表面MHCⅡ类分子相结合。因此SAg发挥作用需有两类细胞:表达TCRβ链的CD4+T细胞和表达MHCⅡ类抗原的辅佐细胞。外源性的SAg主要是葡萄菌、链球菌、支原体等微生物产生的毒素,其中以葡萄球菌肠毒素A(Staphylococcus enterotoxin A,SEA)研究得最多,SEA往往取用特定TCR基因片段Vβ6.9及Vβ22,SEA另一侧与HLA-DR分子β1结构域的α螺旋相结合。
SAg激活CD4+T细胞使之释放IL-2,IFN、TNF等细胞因子,诱导CTL分化为效应细胞,可杀伤对NK、LAK高度抵抗的白血病细胞。
(二)CD3(T3、Leu4)
CD3分子分布于成熟T淋巴细胞表面,至少由γ、δ、ε、ζ、η5种多肽链组成,与T细胞抗原受体非共价连接(图1-3)。CD3单克隆抗体可诱导CD3多肽和TCR共帽形成(co-capping),并诱导T淋巴细胞活化。TCR识别外来抗原与自身MHC分子形成的复合物,CD3对于信号的传递具有重要作用。

图1-3 TCR/CD3结构模式图
T细胞在胸腺发育过程中,CD3γ、δ和ε基因的表达要早于TCRα、β链基因的表达。CD3γ、δ和ε基因产物通过翻译后的修饰形成核心结构,在内织网处,此核心结构与TCRαβ异源双体形成复合物后转移到高尔基氏体,进行N连接的糖基化。ζ-ζ同源双体与TCRαβ/CD3γδεε结合后组成一个完整的复合物TCRαβ/CD3γδεεζζ(少娄分子为TCRαβ/CD3γδεεζη)。最近发现一个分子量为28kDa的ω连或T细胞受体相关蛋白(t cell receptor associated protein,TRAP),可能具有控制TCR/CD3复合物在内织网中装配和转移的功能,但确切的机理尚不明了。ω链不表达于细胞膜表面。
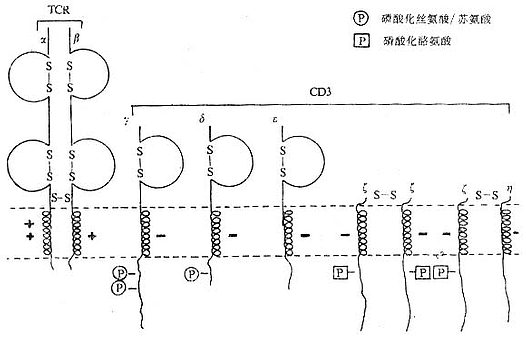
图1-4 TCR/CD3复合物模式图
1.CD3γ、δ和ε链 CD3γ、δ和ε链基因有高度的同源性,在人类位于第11号染色体,小鼠9号染色体,这三种链的基因可能从一个祖先基因通过基因复制而来。CD3γ、δ和ε链在细胞膜外都有一个Ig样结构域(C2),都属于免疫球蛋白超家族,但不存在多态性或可变性,因此不直接参与特异性识别抗原。γ、δ和ε链的穿膜部分含有带负电谷氨酸和/或天冬氨酸残基,这与TCRα、δ链穿膜区中带正电赖氨酸、精氨酸以及β、γ链穿膜区中的赖氨酸相互作用有重要作用。γ、δ和ε链胞浆部分含44到81氨基酸残基,提供了把信息传导到细胞内的条件。γ链分子量为25~28kKa,有2个糖基化点,氨基端89个氨基酸残基为亲水性,组成胞膜外区,穿膜区含27个氨基酸残基,胞浆内区44氨基酸残基,胞浆内113位丝氨酸残基可能是磷酸化位点。δ链分子量为20kDa,含有2个糖基化点,胞浆内126位丝氨酸可能是磷酸化位点。CD3δ链抗体能非特异性地活化T细胞,促进T细胞有丝分裂。ε链分子量为20kDa包括氨基端104亲水氨基酸的胞膜外区,穿膜区26个氨基酸残基,胞浆内区81个氨基酸残基。目前所制备的单克隆抗体中大部分是针对CD3ε链。
2.CD3ζ和η ζ(zeta)和η(eta)链结构相似,而与CD3γ、δ和ε链无同源性。ζ和η链分子量分别为16kDa和21kDa,它们的胞膜外以及穿膜区和结构相似,但有胞浆区不同。胞膜外区很短,只有9个氨基酸残基,含有半胱氨酸,ζζ之间或ζη之间形成二硫键。ζ和η链穿膜部分各有一个带负电的天冬氨酸。ζ和η链胞浆内区分别有113个和155个氨基酸残基,具有多个酪氨酸磷酸化的位点。最近研究证实,CD3ζ链可能与NK细胞上Fcγ受体相连。此外,ζ链与FcεRⅠγ亚单位有很高的同源性。
CD3γ、δ和ε链是单链,而ζ则以ζ-ζ同源双体存在于80~90%T细胞中,有10~20%T细胞则以ζ-η异源双体存在。因此最常见的TCR/CD3复合物的组成形式是TCRαβ/CD3γδεεζζ。
在体外,抗CD3McAb可促进T细胞表达IL-2R,产生IL-2、TNF-α、TNE-β、IFN-γ和IL-4等多种细胞因子,诱导非MHC限制的细胞毒作用,增强T细胞、LAK和NK细胞的杀伤肿瘤作用。
1986年美国FDA已批准应用小鼠抗CD3 McAb治疗急性肾移植排斥反应。CD3McAb治疗心、肝移植排斥反应已完成Ⅲ期临床试验,正申请投放市场。
(三)CD4
CD4和CD8分子分别与MHCⅡ类和Ⅰ类抗原结合,不仅可增强T淋巴细胞与APC或靶细胞结合的程度,而且与刺激信号的传递有关。CD4阳性细胞是MHCⅡ类抗原限制的细胞群,CD8阳性细胞是MHCⅠ类抗原限制的细胞群。有关CD4和CD8抗原在胸腺细胞分化过程中的变化以及CD4、CD8T细胞亚群见第七章“淋巴细胞群及其亚群”。
1.CD4分子的结构 为细胞膜表面单链糖蛋白,人CD4分子由458个氨基酸残基组成,包括信号肽23氨基酸残基,胞膜外区374个氨基酸残基,含2个糖基化点,穿膜区21氨基酸残基,胞浆内区含有40氨基酸残基。胞膜外区具有4个IgV样结构域,属免疫球蛋白超家族成员。
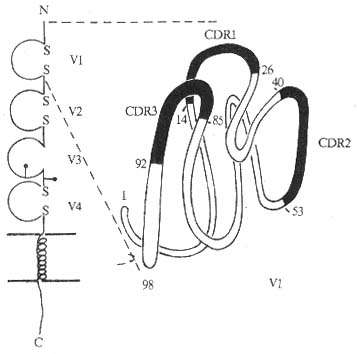
图1-5 CD4分子结构模式图
第一个V样区与Igκ链的V区有很高同源性,有3个互补决定区(complementarity-determining region, CDR)。其余3个V样区功能区与Poly IgR的同源性最接近,其中第2和4个V样区中两个半胱氨酸的距离分别为28和42个氨基酸残基,第3个V样区无二硫键。CD4跨膜区与MHCⅡ类分子β链的跨膜区高度同源。编码人CD4基因位于第2号染色体,小鼠第6号染色体,小鼠CD4分子的分子量为55kDa,由457个氨基酸残基组成,信号肽有22个氨基酸残基,N端功能区110个,胞膜外还有一个长序列(long sequence)的区域,含262氨基酸残基,有4个糖基化点,穿膜区25氨基酸残基,胞浆内区含38个氨基酸残基。人和小鼠CD4分子约有55%序列相同,尤以胞浆内区为显。在胞浆部位有3个丝氨酸残基,可能作为PKC磷酸化的底物。CD4胞浆部分功能区高度的保守性表明这一区域的功能是重要的。
2.CD4分子的分布 分布于部分T淋巴细胞和胸腺细胞表面,也发现于某些B淋巴细胞、EBV转化和B细胞、单核吞噬细胞和脑细胞。在人类,OKT4和Leu3McAb可检测CD4抗原。小鼠L3T4是人OKT4的类同物。
3.CD4分子的功能 在成熟的胸腺细胞、外周血和周围淋巴器官中,CD4阳性细胞一般为辅助性T淋巴细胞诱导细胞/抑制性T淋巴细胞诱导细胞(helper inducer/suppressor inducer)。
(1)作为细胞与细胞之间的粘附分子:CD4第1、2功能区与MHCⅡ类分子的非多态部分结合以稳定MHCⅡ类分子限制的T细胞与带有MHCⅡ类分子与抗原复合物的APC细胞相互作用。抗CD4McAb可封闭T细胞的辅助活性。
(2)转导信号:CD4分子胞浆区与蛋白酪氨酸激酶p56lck相联,对T细胞信号的转导起重要作用(详见第八章)。
CD4分子胞膜外第1个结构区域是HIV外壳蛋白gp120的识别部位,其中CDR2与gp120结合的亲合力最高,CDR3可能与HIV感染靶细胞膜融合有关。可溶性 gp120结合到CD4的反应可被下列试剂所阻断:(1)针对CD4V1区中CDR2、CDR3的McAb;(2)CDR2、CDR3肽段;(3)可溶性CD4 V1肽段;(4)抗gp120抗体。HIV感染机体可引起选择性CD4+细胞的数量减少和功能降低,主要通过以下不同的机理:(1)HIVgp120与T细胞表面CD4分子结合后通过病毒芽生破坏细胞膜,在感染细胞浆内产生大量非整合的病毒RNA直接损伤细胞膜、干扰细胞代谢,影响CD4分子在细胞膜上的表达以及形成短命的合胞体;(2)阻断CD4+T细胞与Mψ细胞表面MHCⅡ类抗原的结合,影响Th细胞对抗原的识别过程;(3)产生抗体损伤CD4细胞,机体产生抗gp120或其他HIV成份的抗体,通过激活补体或ADCC效应损伤CD4阳性细胞;(4)特异性CTL也可通过识别CD4细胞表面的gp120分子而杀伤CD4阳性细胞。最近发现,CD26可能是HIV的另一类受体。
应用基因工程生产的重组可溶性CD4(rsCD4)治疗ARC(AIDsrelated complexes)、艾滋病正在进行Ⅱ期临床试验;抗CD4McAb(Leu3a)也已开始治疗HIV感染的I期临床试验。此外,应用CD4-IgG、CD4-PE(绿脓杆菌外毒素)、CD4-RA(蓖麻毒毒A)等杂交分子杀伤HIV感染的T细胞,作为抗爱滋病的新药也已进入临床验证。1991年美国风湿病学年会上报道了用抗CD4McAb治疗类风湿性关节炎(RA),经治疗后临床症状明显改善,PBMC中CD4阳性细胞的比例和CD4抗原密度明显下降,血清可溶性CD4(sCD4)水平明显升高,血沉、CRP、RF、和总免疫球蛋白水平明显降低。抗CD4嵌合抗体(Centocor公司)治疗类风湿性关节炎、多发性硬化症也已进入Ⅱ期临床验证。抗CD4McAb(Ortho Biotech公司)预防器官移植排斥反应已开始临床验证。
(四)CD8
1.CD8分子的结构 CD8分子是由α、β两条多肽链组成的穿膜糖蛋白,α链分子量34kDa,相当于小鼠的Lyt-2;β链30kDa,相当于小鼠的Lyt-3。每条链各包括1个IgV样结构域、连接肽、穿膜区和胞浆区。α和β链在连接肽处有二硫键相连。部分CD8分子是由同源α链双体(α/α)组成,如在CD8阳性的TCRγδT细胞表面。有报道胸腺细胞上的CD8可能为四聚体。CD8α和β链IgV样区约含110氨基酸残基,与Igκ、λ轻链的V区有30~35%同源性,与V有20~22%同源性,与TCr Vα和Vβ有24%同源性。编码CD8α、β链的基因属Ig基因超家族成员,与编码Igκ链基因密切连锁,定位于第2号染色体,表达前不需要重排。编码小鼠Lyt-2和Lyt -3基因定位于第6号染色体,各有二个等位基Ly2a、Ly2b和Ly3a、Ly3b,分别编码Lyt-2.1、Lyt-2.2和Lyt-3.1和Lyt-3.2。
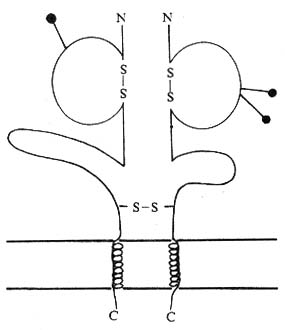
图1-6 CD8分子结构模式图
2.CD8分子的分布 分布于部分T淋巴细胞和胸腺细胞。在异基因骨髓移植病人中可出现TCRγδCD8α/α表型的T细胞。NK细胞表面的CD8分子为α/α二聚体。
最近发现,通过细胞分泌或/和胞膜外分子脱落的机制,在血清中存在着可溶性CD8分子(sCD8)。白血病、何杰金氏病、艾滋病、急性传染性单核细胞增多症、再生障碍性贫血、同种异体移植、类风湿性关节炎和全身性红斑狼疮等患者血清中sCD8水平增高,其升高的水平与疾病的严重程度、病情变化、治疗反应以及预后有较密切的关系。
3.CD8分子的功能
(1)作为细胞与细胞间的附粘分子MHCⅠ类抗原是CD8分子的配体。CD8分子与MHCⅠ类分子结合可以稳定MHCⅠ类分子限制的T细胞(主要是CTL)与带有MHCⅠ类分子与抗原复合物的靶细胞结合。CD8阳性细胞为抑制性T淋巴细胞/杀伤性T淋巴细胞(suppressor T lymphocyte/cytotoxic Tlymphocyte,Ts/Tc)。T8、Leu2 McAb识别CD8α链,可封闭Tc的活性。
(2)转导信号,目前发现CD8也与蛋白酪氨酸激酶p56lck相关。在T细胞增殖和分化的信号转导中起重要作用。
抗CD8McAb可预防骨髓移植时移植物抗宿主反应,美国Becton-Dicknson公司生产Leu2已进入Ⅱ期临床验证。
(五)CD2
1.CD2分子的结构和分布 CD2分子又称T11、绵羊红细胞受体(ER)、淋巴细胞功能相关抗原2(LFA-2)和Leu5,是人T淋巴细胞表面的单链糖蛋白,分子量50kDa,CD2基因定位于第1号染色体,属免疫球蛋白基因超家族。编码351氨基酸残基,包括先导序列24氨基酸残基,2个胞外功能区(C2)共185氨基酸残基,有3个糖基化位点,穿膜区和胞浆部分分别为26和116个氨基酸残基,胞浆区富含肺氨酸和碱性氨基酸,胞浆区与活化信号的传递可能有关。在DNA水平上人和小鼠CD2有51%同源性。CD2分子分布于95%的T细胞、50~70%胸腺细胞和大颗粒淋巴细胞(LGL/NK)。
2.CD2分子的功能
(1)粘附功能:CD2分子的配体是LFA-3,后者分布于多种细胞表面。CD2阳性T细胞可结合含有纯化LFA-3的脂质体。CD2分子的功能主要是通过抗CD2McAb对淋巴细胞功能的影响和基因转染技术来研究的。用抗CD2McAb可(1)抑制lectin、同种异体抗原、可溶性抗原等诱导T细胞的增殖反应;(2)抑制T淋巴细胞IL-2合成和分泌;(3)抑制CTL效应相杀伤功能和NK细胞的杀伤活性。CTL与靶细胞之间的抗原非特异性粘附有多种途径,如CD2与LFA-3结合,LFA-1与ICAM-1结合。用抗CD2和抗LFA-1两种抗体,或抗LFA-1和抗LFA-3两种抗体可完全抑制抗原非特异性粘附作用,而抗CD2与抗LFA-3两种抗体只能部分抑制这种粘附作用。CD2与LFA-3之间的粘附功能对于T淋巴细胞TCR识别外来抗原与APC细胞表面MHC抗原复合物、肿瘤抗原、病毒感染靶细胞以及同种异体抗原均有重要的辅助作用。最近研究表明,CD48和CD59也是CD2的配体,参与T细胞的粘附和细胞间的相互作用。
(2)T细胞旁路激活途径(the alternative pathway of T cellactivation): Reinherz成功地制备了识别了CD2三个不同表位的McAbs:(1)T111与LFA-3(CD58)的结合有关,由于绵羊红细胞SRBC表达CD58的同源物,T111可与SRBC结合形成E花环,抗T111McAb可抑制E花环形成;(2)T112与CD58结合无关,抗T112McAb可诱导T113表位出现;(3)T113是活化T细胞表达的表位(已命名为CD2R),在诱导T113表位出现的过程中,加入蛋白质合成抑制剂T113仍能表达,表明T113出现并非是一种新合成的蛋白质,而是由于活化后构型变化暴露出来的表位。同时加入抗T112和T113McAbs可活化T淋巴细胞,促进MHCⅡ类抗原和IL-2受体的表达,并在IL-2受体的表达,并在IL-2存在条件下,活化的T细胞继续增殖,称为T细胞旁路激活途径。T113McAb与表达IL-3的SRBC同T细胞一起孵育,也能刺激T细胞增殖。CD3阴性的胸腺细胞和NK细胞可因CD2而活化,此外,CD2可以与其它膜分子如CD44、CD45的功能有关。
目前关于CD2活化的生理学意义还不十分清楚,CD2阳性胸腺细胞可与LFA-3阳性的胸腺上皮细胞结合,在胸腺微环境的调节下,可能为早期胸腺细胞的活化提供信号,并与胸腺细胞的增殖和分化有关。
(六)CD58(LFA-3)
1.LFA-3分子的结构 淋巴细胞功能相关抗原-3(lymphocyte functionassociated antigen-3,LFA-3)(CD58)是细胞表面糖蛋白,分子量55~70kDa,属免疫球蛋白超家族成员,与CD2分子高度同源,胞膜外区有2个Ig超家族C2样区。CD2和CD58基因都定位于1号染色体,并密切连锁,可能是从同一个基因复制而来。CD2和CD58的结合是属于嗜同种的相互作用(homophilic interaction)。
2.LFA-3分子的分布 分布于T细胞、B细胞、单核细胞、上皮细胞、内皮细胞、结缔组织、成纤维细胞、中性粒细胞和血小板表面。
3.LFA-3分子的功能 LFA-3分子的功能主要是通过应用相应的McAb而得知,抗LFA-3抗体可(1)抑制效应相CTL的活性,可能与抑制CTL与靶细胞之间的粘附有关;(2)抑制lectin、同种异体抗原诱导的T细胞增殖反应。在EB病毒感染的Burkitt氏淋巴瘤细胞株中,发现有的肿瘤细胞由于缺乏LFA-3的表达而抵抗CTL的杀伤作用,提示LFA-3缺损的肿瘤细胞可能与逃逸机体的免疫监视有关。发作性夜间血红素尿症(paroxysmalmocturnal hemoglobinuria,PNH)患者红细胞表面缺乏LFA-3、DAF和FcγRⅢ。
LFA-3以两种形式存在于细胞膜表面:(1)穿膜形式,胞膜外区、穿膜区和胞浆区分别为188、23和12个氨基酸;(2)GPI“锚”形式,如EBV转化的JYB细胞系细胞表面LFA-3分子具有穿膜和GPI“锚”两种形式。
[GPI“锚”]糖基磷脂酰肌醇(glycosylphosphatidylinsitol,GPI)是质膜的组成成分,GPI骨架上的乙醇胺通过酰胺键固定于蛋白质的羧基端,成为蛋白质定位于细胞膜上的锚,这种结合到GPI的蛋白质即称为GPI锚蛋白。去污剂溶解脂质双层可获得结合在GPI上的蛋白质,磷脂酶C(PLC)可从细胞表面将糖一肌醇磷脂锚连的蛋白质释放出来,磷脂酰肌醇经PLC降解后所产生的三磷酸肌醇(IP3)和二酰基甘油(DG)是重要的第二信使。
(七)CD28
1.CD28分子的结构 1980年Hara等首先用单克隆抗体9.3发现了CD28。CD28由两条44kDa多肽借二硫键组成的同源二聚体,分子量为90kDa。成熟的人CD28分子单肽链有202个氨基酸,基中胞膜外区有134氨基酸,属免疫球蛋白超家族成员,有一个IgV样区。人与小鼠CD28分子的同源性为68%。CD28与CTLA-4分子有高度同源,后者主要表达于活化的CTL细胞表面。
2.CD28分子的分布 在外周血淋巴细胞,CD28+细胞占54~86%,其中90%CD4+T细胞和50%CD8+T细胞表达CD28。CD28在CD28+T细胞中表达与功能有一定的关系,CD8+CD28+T细胞表现出MHC限制的细胞毒功能,而CD8+CD28-细胞可抑制抗体产生以及同种异体抗原所诱导的细胞增殖效应。此外,浆细胞瘤及部分活化B细胞也可表达CD28。
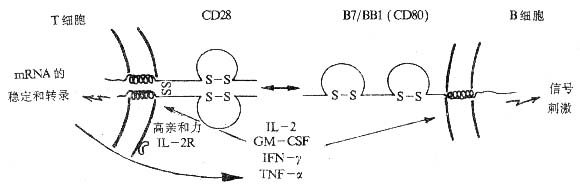
图1-7 CD28与CD80分子相互作用示意图
3.CD28分子的功能
(1)T细胞活化的辅助信号:抗CD28可加强PHA、ConA、抗CD2McAb、抗CD3McAb等增殖、活化的效应,增加IL-2、IL-3、TNF-α、IFN-β、GM-γ、GM-CSF等细胞因子产生,激活CTL细胞。此外,抗CD28McAb可诱导T细胞IL-13mRNA的表达。
(2)CD28的天然配体是B细胞活化抗原B7/BB1(CD80),CD28与CD80的结合是T-B细胞相互协作的主要方式,并刺激B细胞活化。
二、与B细胞识别、粘附、活化有关的CD分子
(一)BCR复合物
B细胞抗原受体(B cellreceptor,BCR)复合物至少由四种不同的多肽链组成,抗原结合部位是由重链和轻链构成的膜表面Ig的四链结构,此外,在BCR中还含有Igα和Igβ两种多肽链,最近在白细胞分化抗原国际专题讨论会中分别命名为CD79a和CD79b。在人类B细胞,与mIgM相关的Igα和Igβ分别为47kDa和37kDa糖蛋白,属于免疫球蛋白超家族成员,编码Igα和Igβ的基因分别称为mb-1和B29。Igα和Igβ胞膜外区氨基端处均有一个Ig样结构域。Igα和Igβ均可作为蛋白酪氨酸激酶的底物,可能与BCR信号转导有关,因为mIgM和mIgD胞浆区只有3个氨基酸(KVK),不可能单独把胞膜外的刺激信号传递到细胞内。Igα和Igβ胞浆部分尾部有6个保守的氨基酸残基,可能以磷酸化形式与胞浆中不同酶中存在的SH2(src-homology2)结构域结合。
(二)CD19
CD19是一种属于Ig超家族成员、分子量为95kDa的穿膜糖蛋白,分布于B细胞表面,其相应的生理性配体尚不清楚。CD19与B细胞活化和信号的转导有关:(1)CD19单克隆抗体可诱导胞浆内多种底物迅速发生磷酸化;(2)CD19胞浆区可被一种丝氨酸激酶催化而发生磷酸化;(3)CD19胞浆区与src激酶家族Lyn稳定的结合。最近提出一个B细胞最佳信号放大的双重抗原结合模型,这个模式认为B淋巴细胞BCR/Igα、Igβ与抗原结合后,使CD19与CD21相互接近形成复合物,外来抗原及包裹抗原的C3dg分别被BCR和CD21所结合,后者激活CD19/CD21复合物中与CD19紧密结合的src家族Lyn,而使CD19分子胞浆内酪氨酸发生磷酸化,有关信号转导过程参见第八章“淋巴细胞活化过程中信号转导的分子基础”。
(三)CD21
CD21又称2型补体受体(complementreceptor type 2,CR2)和EB病毒受体,是补体激活调节剂家族的一员。
[补体激活调节剂]补体激活调节剂(regulatorsof complement activation,RCA)家族包括2个血浆蛋白H因子和C4bp以及4个膜蛋白CR1、CR2、DAF和MCP。RCA的特点是:(1)含有60~70个氨基酸组成的短同源重复顺序(short consensus repeat ,SCR);(2)结合补体活化裂解片段C3b和C4b;(3)基因定位于染色体1q32处,其基因以MCP-CR1-CR2-DAF-C4bp形式相连锁。
1.CD21分子的结构 为分子量140kDa的单链糖蛋白,N端在细胞外,胞膜外区1005个氨基酸,疏水跨膜区28个氨基酸,富含碱性氨基酸,胞浆区34个氨基酸。胞膜外区组成15个SCR,每个SCR含有60~70氨基酸,有一个含4个Cys的骨架结构,C1-C4、C2-C3间形成两个二硫键,构成一个SCR球状结构,SCR1-2与C3dg包裹的颗粒和EBV结合有关,胞膜外区有10个(或12个)N-糖键。胞浆区有10个可能磷酸化为位点。
2.CD21的分布 主要分布在成熟的B细胞、淋巴滤泡内树突状细胞、部分T细胞,此外,口咽、鼻咽以及宫颈上皮细胞表达与CD21相关的145kDa分子。
3.CD21的功能
(1)促进B细胞增殖:单独CD21交联并不引起B细胞的增殖,在T细胞或T细胞源性低分子量B细胞生长因子(LMW-BCGF)存在下,或抗μ链抗体激活B细胞增殖时,CD21交联具有强烈的促进作用;聚合的C3dg、UV-EBV、CD21单抗可加强TPA刺激的B细胞增殖;在LMW-BCGF存在下,CD21单抗可刺激B细胞增殖;C3dg包裹的颗粒可引起LPS激活的小鼠B细胞连续增殖;可溶性入C3dg诱导Raji细胞不信赖血清的增殖。上述结果提示外周血B细胞被某些刺激物刺激后,CD21介导增殖信号促进B细胞进入细胞周期。最近发现CD23是CD21的一个新的配体。sCD23在体外促进IgE生成,并有BCGF样作用,CD23分子能与CD21结合,推测CD21就是生长因子BCGF的受体。
(2)CD21介导EBV转化B细胞:EBV与CD21(CR2)结合后激活磷脂酶C,水解磷脂酰肌醇,胞浆内Ca2+和二酰基甘油水平增加,并活化钙调蛋白和PKC。转化性EBV(如B95-8)感染B细胞后,编码反式激活蛋白(transactivator)EBNA2、LMP激活CD21和CD23基因,导致B细胞持续高水平表达CD21和CD23,CD23脱落后形成sCD23是自分泌的BCGF,与CD21结合并活化CD21分子的酪氨酸蛋白激酶,不断激活PKC,从而导致B细胞的转化和增殖。
(3)CD21参与免疫记忆:病原微生物或蛋白质抗原上覆盖有C3dg时,可与淋巴滤泡内树突状细胞表面CD21结合,在诱导免疫记忆过程中起重要作用。
(4)参与补体的活化:CD21参与补体替代途径的启动以及C3b的固定,且在C3bi裂解为C3dg过程中为丝氨酸蛋白酶Ⅰ因子的辅因子。
(四)CD80(B7/BB1)
CD80cDNA已克隆成功,成熟的CD80分子由262氨斟酸组成,胞膜外区216个氨基酸,穿膜区27氨基酸,胞浆区19氨基酸。B7和BB1分子量分别为44kDa和46kDa,其分子量的差异是由同一核心多肽糖基化程度不同所造成。CD80属免疫球蛋白超家族成员,是B细胞活化抗原,IFN-γ活化的单核细胞也表达CD80。CD28是CD80的配体,CD28与CD80的结合可同时引起T细胞和B细胞的活化,在T、B细胞协作中发挥重要作用。
三、免疫球蛋白Fc段受体
Ig根据其重链抗原性的差异分为IgM、IgG、IgA、IgD和IgE五类,各类Ig的不同功能主要与其结构有关。机体内许多细胞表面具有不同类Ig Fc的受体,通过Fc受体与Ig Fc的结合,参与Ig介导的生理功能或病理损伤过程。目前已鉴定明确属于CD抗原的Fc受体有FcγR、FcαR、FcεR。
(一)FcγR(CD64、CD32、CD16)
1.FcγR的结构和分布 FcγR可分为FcγRⅠ、FcγRⅡ和FcγRⅢ三类,它们的结构和分布有所不同。
(1)FcγRⅠ(CD64):70kDa穿膜糖蛋白,属Ig超家族成员,胞膜外区有3个C2结构,基因染色体定位于1q23~24。识别CD64的代表性McAb有McAb22、McAb32.2、197和10.1等FcγRⅠ是高亲合力受体,Kd值为10-8~10-9M,主要与人的单体IgG1、IgG3以及小鼠IgG2a和IgG3结合。与人IgG4结合的亲合力明确降低,与IgG2则无结合能力。FcγRⅠ主要分布于单核细胞、巨噬细胞、中性粒细胞等,但表达水平各不相同。FcγRⅠ位点数:15000~40000/每个单核细胞,>50000/巨噬细胞,<1000/新鲜中性粒细胞。IFN-γ可刺激单核细胞、巨噬细胞和中性粒细胞表达FcγRⅠ水平增加5~10倍,G-CSF也有这种促进作用。
(2)FcγRⅡ(CD32):40kDa穿膜糖蛋白,属于Ig超家族成员,胞膜外区有2个C2结构,基因染色体定位于1q23~24。识别CD32的代表性McAb有CIkM5、IV·3、KuFc79和41H16等。FcγRⅡ与单体人IgG1,IgG3、IgG4结合为低亲合力,Kd5×10-7M。FcγRⅡ表达于除红细胞外的其它血细胞,分子数目:20000~40000/每个细胞。根据DNA序列和功能不同,FcγRⅡ可分为三种形式,不同形式FcγRⅡ的差别主要在于胞浆区的结构不同。
(3)FcγRⅢ(CD16):50~70kDa糖蛋白,属Ig超家族成员,有2个C2结构,基因染色体位于1q23~24。识别CD16代表性的McAb有HUNK2、Leu11、3G8、Gran1和B73.1等。FcγRⅢ结合人IgG、IgG3,为低亲和力受体。FcγRⅢ有FcγRⅢA和FcγRⅢB两种异型:①FcγRⅢA,穿膜结构,主要分布于巨噬细胞、NK细胞和嗜酸性粒细胞,巨噬细胞表达高水平FcγRⅢA,而单核细胞表达水平较低。FcγRⅢA与二硫键连接的CD3ζ或FcεRⅠγ链双体相关,巨噬细胞上FcγRⅢA与CD3复合体γ链相关,NK/LGL上FcγRⅢA则与ζ链相关。TGF-β促进培养的单核细胞表达FcγRⅢA。②FcγRⅢB,通过GPI“锚”在中性粒细胞表面,每个人中性粒细胞表达10万~20万血液中可溶性的FcγRⅢ主要来自这种形式,中性粒细胞激活剂短时间处理后可明显降低FcγRⅢB的表达水平,可能与通过激活内源性蛋白酶切除GPI连接分子有关。

图1-8 FcγR、FcαR和FcεR结构示意图
2.FcγR的功能 FcγR的功能主要是通过髓样细胞和NK细胞来发挥的。
(1)单核-巨噬细胞:FcγRⅠ、Ⅱ和Ⅲ均可介导人单核细胞ADCC来杀伤肿瘤等靶细胞,这种ADCC效应为Mg2+依赖,并需LFA-1等粘附分子参与。IFN-γ可促进单核细胞FcγRⅠ介导的杀伤作用。单核-吞噬细胞可通过FcγRⅠ、Ⅱ、Ⅲ发挥调理吞噬和清除免疫复合物的作用。
(2)中性粒细胞:新鲜分离的中性粒细胞不能通过FcγR溶解靶细胞,但在IFN-γ刺激下可通过FcγRⅠ和FcγRⅡ介导杀伤作用,对于FcγRⅠ,IFN-γ主要是诱导其表达水平升高,而对FcγRⅡ表达水平并未见改变,可能是通过对杀伤机理的调节。GM-CSF也能通过FcγRⅡ明确增加中性粒细胞的杀伤水平。GPI连接的FcγRⅢB并不能介导中性粒细胞杀伤肿瘤的作用。活化中性粒细胞通过FcγRⅠ、Ⅱ发挥调理吞噬和清除免疫复合物的作用。
(3)嗜酸性粒细胞:未刺激的嗜酸性粒细胞没有杀伤作用,GM-CSF、TNF和IL-5等是嗜酸性粒细胞发挥ADCC效应的有效激活剂,在杀伤寄生虫和抗肿瘤中有重要作用。GM-CSF对嗜酸性粒细胞的激活作用主要是通过FcγRⅡ介导的。
(4)NK细胞:通过FcγRⅢA介导ADCC杀伤肿瘤细胞等靶细胞,IL-2和IFN-γ可明显提高NK细胞的杀伤活性,但并不明显改变FcγRⅢA的表达水平。
(二)FcαR(CD89)
FcαR(CD89)为分子量60kDa穿膜糖蛋白,胞膜外区206氨基酸,穿膜区19氨基酸,胞浆区为41氨基酸,属Ig超家族成员,胞膜外有2个C2结构域,为中亲和力受体,主要表达于单核细胞、巨噬细胞、中性粒细胞等,介导吞噬、ADCC以及炎症介质的释放。中性粒细胞表面FcαR可结合血清型和分泌型IgA1和IgA2,亲和力约为Kd5×10-7M。热或化学物质凝集的IgA可刺激中性粒细胞脱颗粒。
(三)FcεR(FcεRⅠ、FcεRⅡ)
1.FcεR的结构和分布 FcεR可分为FcεRⅠ和FcεRⅡ两类,其结构、分布及介导的生物学作用有所不同。
(1)FcεRⅠ:为高亲和力受体,Kd10-9~10-10M,由α、β、γ-γ四条链组成。其中α链含222个氨基酸残基,分子量为25kDa,胞膜外区属Ig超家族结构,2个C2区,与FcγRⅡ和FcγRⅢ高度同源,胞膜外区与IgECε2/Cε3结合;穿膜区20左右氨基酸中含有与FcγRⅢA相同的8个氨基酸残基;胞浆区的31个氨基酸结构较为独特。β链含243个氨基酸残基,分子量为33kDa,有四个穿膜部分,N端与C端都位于胞浆内,β链可能把α链和γ-γ链连在一起。两条γ链由二硫键连接组成同源二聚体,每条链62个氨基酸,分子量为8kDa,胞膜外区只有5个氨基酸残基,γ链与CD35高度同源,γ链与FcεRⅡ表达的稳定性和信号的转导有关。NK细胞表面FcγRⅢA(CD16)可能与CD3ζ或FcεRⅠγ链相连,提示FcεRⅠγ链与CD3复合物中ζ的结构和功能的相似性。FcεRⅠ主要分布于嗜碱性粒细胞和肥大细胞。
(2)FcεRⅡ(CD23):低亲和力受体,分子量45kDa,单链穿膜糖蛋白,Ⅱ型跨膜蛋白,属C型植物血凝素家族成员。CD23含有321个氨基酸,N端在胞膜内,1~23位氨基酸组成胞浆尾,24~43位氨基酸为疏水跨膜区,靠C端胞膜外区由277个氨基酸组成,有一个糖基化点,82、102、125、150位氨基酸残基为蛋白水解酶敏感位点,凝集素同源区位于C端163Cys至282Cys之间,该同源区共含6个Cys。88至116位氨基酸之间有一个亮氨酸拉链结构,参与CD23分子同源二聚体的形成。CD23分子靠胞膜外C端裂解的不同片段14kDa、25kDa和33~37kDa片段均称为IgE结合因子(IgE-binding factor IgE-BF)。FcεRⅡ可在蛋白水解酶裂解后形成可溶性CD23分子(sCD23)即IgE-BF。CD23mRNA有FcεRⅡamRNA和FcεRⅡbmRNA两种,它们所翻译的CD23分子仅在胞浆区有7个氨基酸残基的差别(图1-9)。FcεRⅡa仅B细胞表达,并易降解为sCD23;FcεRⅡb表达于B细胞、T细胞、嗜酸性粒细胞、血小板、单核细胞、巨噬细胞、树突状细胞、朗罕氏细胞、含有EBV基因组的鼻咽癌细胞、髓样细胞系如U937等,主要以膜分子形式存在,IgECε3与FcεRⅡ结合有关。IL-4可诱导正常B细胞、单核细胞、嗜酸性粒细胞转录FcεRⅡbmRNA,促进CD23的合成与表达,EBV核蛋白EBNA2对CD23表达及sCD23的释放也有促进作用。而IFN-γ、TGF-β、PGE2、糖皮质激素等对B细胞表达CD23和释放sCD23则起抑制作用。
2.FcεR的功能
(1)FcεRⅠ:嗜碱性粒细胞和肥大细胞具有高亲和力FcεRⅠ,每个细胞表面约有10万个,当相应变应原与啫碱性粒细胞、肥大细胞表面IgE/FcεRⅠ复合物结合后通过交联使磷酸肌醇水解,胞浆Ca2+浓度增加,使细胞脱颗粒,合成和释放组织胺、LT、PAF等多种介质,介导Ⅰ型速发型超敏反应。
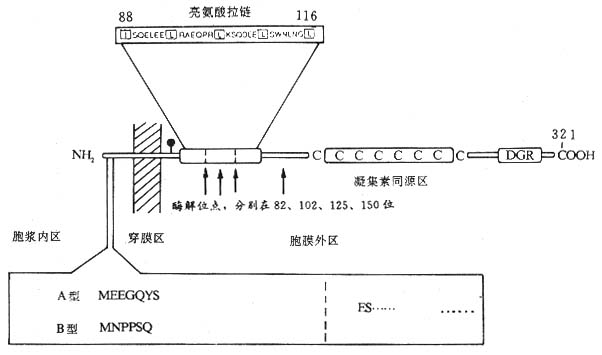
图1-9 CD23分子结构模式图
(2)FcεRⅡ:FcεRⅡ为B细胞发化激活抗原,变态反应性疾病患者PBMC中CD23密度明显增加,血清IgE-BP(sCD23)升高。sCD23具有B细胞生长因子(bcell growth. factor, BCGF)活性,故又称B细胞来源的B细胞生长因子(B-BCGF)。sCD23的这种生长因子作用可能是作为配体与受体CD21(CR2)结合后介导的,CD23分子通过可结合碳水化合物的凝集素同源区与CD21糖链结合。此外,sCD23通过亮氨酸拉链结构,引起B细胞膜CD21分子交联,促进B细胞生长。sCD23对膜CD23有正反馈作用,促进B细胞的分化和IgE的产生,并与IL-4有协同作用,此外,FcεRⅡ还可介导IgE依赖的ADCC和吞噬作用。CD23与B淋巴细胞的转化及恶变有关,EBV转化B细胞后只有B细胞表达CD23才能建立永生化的细胞系,可能与EBV核蛋白的EBNA2有关,EBNA2结合于FcεRⅡa基因起始部位-275~-89的一个189bp DNA片段结合,反式作用于FcεRⅡa基因启动子,并诱导B细胞高表达CD23,sCD23引起膜CD21分子交联,形成一种自分泌生长机制。慢性B淋巴细胞白血病(B-CLL)患者B细胞表达CD23增加,患者血清中sCD23水平明显升高。
关于IgM受体、IgD受体也有报道,前者主要表达于B细胞,后者表达于成熟B细胞。这两种Ig Fc受体尚未归入CD的编号,故在本节从略。此外与多聚IgA和IgM跨膜转运至胞外分泌液中有关的多聚Ig受体(Poly-IgR),通过与多聚Ig中的J链结合而介导Ig转运功能,PolyIgR属免疫球蛋白超家族成员。
(金伯泉)
-
《细胞和分子免疫学》 中的相关章节:
第一章 白细胞分化抗原
第一节 白细胞分化抗原的分类
第二节 白细胞分化抗原的应用(当前页)
参考文献
第二章 粘附分子
第一节 粘附分子的种类和结构
第二节 粘附分子的表达的调节
第三节 粘附分子的功能
第四节 可溶性粘附分子
参考文献
……