第五章 传出神经系统药理概论
传出神经系统包括植物神经系统和运动神经系统。植物神经系统(vegetativenervous system)也称自主神经系统(autonomicnervous system),主要支配心肌、平滑肌和腺体等效应器;运动神经系统则支配骨骼肌。
自主神经系统排除传出神经外,尚包括内脏传入感觉神经,然对后者的生理和药理研究不多。国外文献沿用自主神经药理一词,实际上主要指传出而不包括传入神经药理。此外,自主神经系统不应包括运动神经,但运动神经系统的递质和受体与植物神经系统同属一个体系,而传出神经系统药理一词自可将这两类都概括进来。因此,我国沿用传出神经系统药理一词较为合理。
植物神经自中枢神经系统发出后,都要经过神经节中的突触,更换神经元,然后才达到效应器(effector)。因此,植物神经有节前纤维和节后纤维之分(图5-1,5-2)。
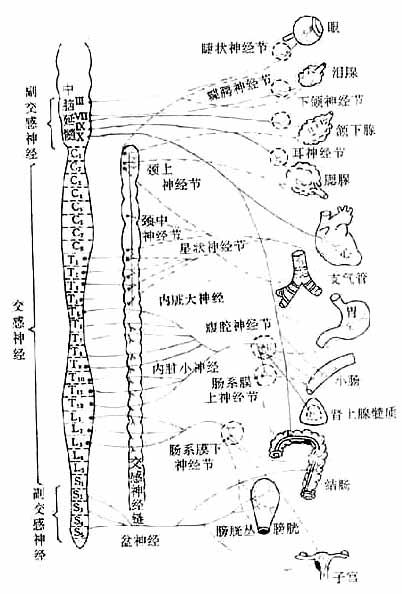
图5-1 植物神经系统分布示意图
神经纤维:蓝色:胆碱能神经 实线:节前纤维
红色:去甲肾上腺素能神经 虚线:节后纤维
运动神经自中枢发出后,中途不更换神经元,直接达到骨骼肌,因此无节前和节后纤维之分。
一、传出神经系统的递质及受体
当神经冲动达到神经末梢时,在突触部位从末梢释放出化学传递物,称为递质(transmitter)。通过递质作用于次一级神经元或效应器的受体(receptor),发生效应,从而完成神经冲动的传递过程。作用于传出神经系统的药物主要是在突触部位影响递质或受体而发挥作用。
(一)传出神经系统的递质
1.递质学说的发展 1921年Loewi通过动物实验证明递质的存在。实验是用两个离体蛙心进行,当刺激甲蛙心的迷走交感神经干以引起迷走神经兴奋时,甲蛙心受到抑制,这时将甲蛙心的灌注液注入乙蛙心,则乙蛙心也表现出抑制。这就说明甲蛙心迷走神经兴奋时,必定释出一种抑制性物质,才能使乙蛙心也受到抑制。后来证明这种物质就是乙酰胆碱。此后相继发现神经节中的节前纤维末梢和运动神经末梢兴奋时,都能释放乙酰胆碱。本世纪四十年代,通过von Euler的工作证明交感神经节后纤维的神经递质是去甲肾上腺素。至此,传出神经系统的化学传递学说才臻完善。
2.传出神经突触的超微结构突触中神经末梢与效应器细胞或次一级神经元间有一定的间隙,称突触间隙。传出神经末梢邻近间隙的细胞膜称为突触前膜;效应器或次一级神经元邻近间隙的细胞膜称为突触后膜。在运动神经与骨骼肌的接头(也称终板),这个间隙约为15~20nm;终板的突触后膜有许多皱褶,其中聚集着胆碱酯酶,能迅速水解已释放的乙酰胆碱。在神经末梢内靠近突触前膜处,聚集着很多直径为20~50nm的囊泡(vesicle),囊泡内含有大量递质乙酰胆碱。
交感神经末梢分成许多细微的神经纤维,分布于平滑肌细胞之间。这些细微神经纤维都有稀疏串珠状的膨胀部分,称为膨体(varicosity)。膨体中含有线粒体和囊泡等亚细胞结构,一个膨体内囊泡的数目约在1000个左右。囊泡内含有高浓度的去甲肾上腺素。
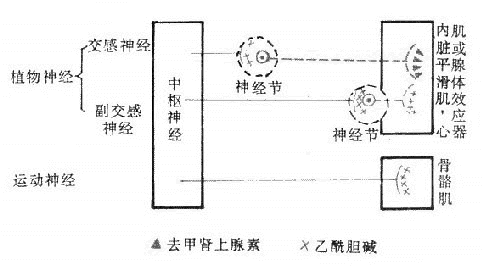
图5-2 传出神经分类模式图
蓝色:胆碱能神经 实线:节前纤维
红色:去甲肾上腺素能神经 虚线:节后纤维
去甲肾上腺素能神经内的囊泡有大小之分,大囊泡在神经节细胞内形成,以每小时数mm的速度沿轴突向末梢运行;小囊泡主要在神经末梢形成。运行到末梢的还有合成去甲肾上腺素所必需的酶,如酪氨酸羟化酶、多巴脱羧酶和多巴胺β-羟化酶等,后者存在于囊泡内;前二者存在于胞质液中。
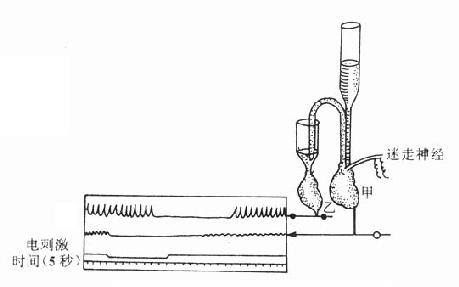
图5-3 证明迷走神经兴奋时释放递质的双蛙心实验
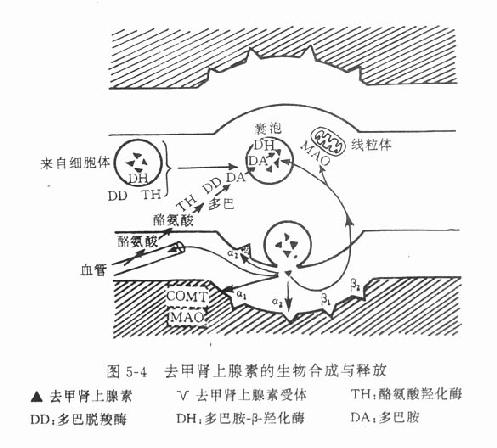
3.递质的生物合成与贮存 去甲肾上腺素的生物合成在去甲肾上腺素能神经细胞体内和轴突中即开始进行,不过在此含量较少,愈到神经末梢,含量愈多,末梢内的含量约为细胞体内的3~300倍。酪氨酸从血液进入神经元后,在酪氨酸羟化酶催化下生成多巴(dopa)再经多巴脱羧酶的催化,脱羧后生成多巴胺(dopamine),后者进入囊泡中,经多巴胺β-羟化酶的催化,转变为去甲肾上腺素。酪氨酸羟化酶的活性较低,反应速度慢,底物要求专一,当胞浆中多巴胺或游离的去甲肾上腺素浓度增高时,对该酶有反馈性抑制作用,反之,当胞浆中多巴胺或去甲肾上腺素浓度降低时,对该酶的抑制作用减弱,催化反应则加速,故这一步骤是去甲肾上腺素生物合成过程的限速因素,是调节去甲肾上腺素生物合成的重要环节。去甲肾上腺素形成后,与ATP的嗜铬颗粒蛋白结合,贮存于囊泡中(图5-4),并可避免被胞质液中的单胺氧化酶(MAO)所破坏。
乙酰胆碱主要在胆碱能神经末梢形成,与其合成有关的酶和辅酶有胆碱乙酰化酶(cholineacetylase)和乙酰辅酶A(acetyl coenzyme A)。前者在细胞体内形成并沿轴突转运至末梢,后者则在末梢线粒体内合成,须先与草酰乙酸缩合成枸橼酸盐,才能穿过线粒体膜进入胞质液中,然后在枸橼酸裂酶的催化下再形成乙酰辅酶A。胆碱乙酰化酶和乙酰辅酶A在胞质液内促进胆碱形成乙酰胆碱。乙酰胆碱形成后,即进入囊泡并与ATP和囊泡蛋白共同贮存于囊泡中(图5-5)。
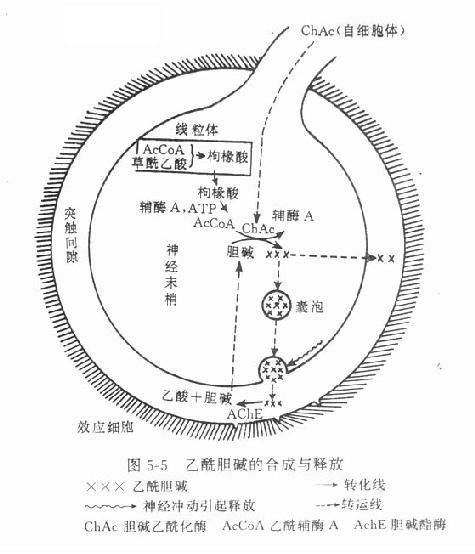
4.递质的释放 现认为当神经冲动到达末梢时,产生除极化,引起Ca2+内流促使靠近突触前膜的一些囊泡的囊泡膜与突触前膜融合,形成裂孔,通过裂孔将囊泡内的递质、ATP和蛋白质等排出至突触间隙,这称为胞裂外排(exocytosis)。每一囊泡约含有1000~50000分子乙酰胆碱或约10000分子去甲肾上腺素。骨骼肌或平滑肌细胞有自发性小终板电位(miniatuse endplate potential)或接头电位,其电位幅度有分极现象;因此提出递质的量子化释放(quantalrelease)概念。每一量子相当于一个囊泡的释放量。由于这种电位幅度很小(0.3~3.0mv),故不会引起动作电位和效应。冲动到达时,可有上百个囊泡同时外排,才引起动作电位和效应。近知嗜铬细胞胞浆中的会合素(synexin),在有Ca2+时,能融合嗜铬颗粒(相当于神经末梢的囊泡)膜与细胞膜。
5.递质作用的消失 乙酰胆碱作用的消失主要是被神经突触部位的胆碱酯酶水解,一般在释放后一至数毫秒之内即被此酶水解而失效。去甲肾上腺素主要靠突触前膜将其摄取入神经末梢内而使作用消失;这种摄取称为摄取1(uptake1)。摄取1是一种主动的转运机制,也称胺泵(amine pump),能逆浓度梯度而摄取内及外源性去甲肾上腺素。其摄取量为释放量的75%~95%,摄取入神经末梢的去甲肾上腺素尚可进一步被摄取入囊泡,贮存起来以供下次的释放。部分未进入囊泡的去甲肾上腺素可被胞质液中线粒体膜上的单胺氧化酶(mono-amineoxidase,MAO)破坏。非神经组织如心肌、平滑肌等也能摄取去甲肾上腺素,称为摄取2。此种摄取之后,即被细胞内的儿茶酚氧位甲基转移酶(catechol-O-methyltransferase,COMT)和MAO所破坏;因此摄取1可称为摄取-贮存型,摄取2可称为摄取-代谢型。此外,尚有小部分去甲肾上腺素释放后从突触间隙扩散到血液中,最后被肝、肾等的COMT和MAO所破坏。
(二)传出神经系统的受体
1.受体的命名 受体的命名常根据能与之选择性地相结合的递质或药物而定。能与乙酰胆碱结合的受体,称为胆碱受体(cholinoceptor)。由于在早期的研究中,发现位于副产感神经节后纤维所支配的效应器细胞膜的胆碱受体对以毒蕈碱为代表的拟胆碱药较为敏感,故这部分受体称为毒蕈碱(muscarine)型胆碱受体(M胆碱受体)。位于神经节细胞膜和骨骼肌细胞膜的胆碱受体对烟碱比较敏感,故这些部位的受体称为烟碱(nicotine)型胆碱受体(N胆碱受体),也可将前者称为N1受体,后者称为N2受体。近年发现M胆碱受体也有M1受体、M2受体和M3受体之分。M1受体主要分布于神经节细胞和腺体细胞,哌仑西平能选择性地阻断之;M2受体似主要分布于心脏;M3受体似主要分布于平滑肌和腺体细胞。哌仑西平对M2和M3受体的亲和力较低,阿托品对三种M受体都能阻断。
能与去甲肾上腺素或肾上腺素结合的受体称为肾上腺素受体(adrenoceptor)。肾上腺素受体又可分为α肾上腺素受体(α受体)和β肾上腺素受体(β受体)。β受体又分为β1受体和β2受体两种亚型,例如心脏的β受体主要为β1受体,支气管和血管平滑肌的β受体主要为β2受体。
70年代发现外源去甲肾上腺素可抑制去甲肾上腺素能神经内[3H]标记去甲肾上腺素的释放(图5-6)。因此认为突触前膜也有α受体,激动时可使递质释放减少,这是一种递质释放的自身调节机制。当时将突触后膜α受体命名为α1受体,突触前膜受体命名为α2受体。后来在许多血管组织突触后膜都发现有不受α1受体阻断药哌唑嗪阻断的α2受体。血管平滑肌突触后膜的α1和α2受体的共存似为普遍现象,可见以突触前、后膜的解剖部位进行α受体分类是不妥的。而以特异的阻断剂和激动剂来区分受体亚型更为合适。
![NA对电刺激所致[3H]NA释放的影响](../../image/lilunshuji/yaolixue/yaolixue058.jpg)
图5-6 NA对电刺激所致[3H]NA释放的影响
猫脾脏灌流,每次刺激共200脉冲,30Hz,
括号内数字为实验次数,竖线为SE(自Bevan,1978)
2.受体按偶联和结构的分类根据第二章对受体的叙述,肾上腺素受体和M胆碱受体属于G-蛋白偶联受体,N胆碱受体属于配体门控通道型受体。近年由于单克隆抗体和DNA重组等技术的应用,许多受体的一级结构得以阐明。现已知G-蛋白偶联受体一级结构的特点是都有7个跨膜区段,以β2受体为例,自图5-7可见,每个跨膜区段有20余个氨基酸残基组成的亲脂性螺旋结构。其伸出细胞膜外的N端较短,伸入细胞内的C端较长。处于β受体第3跨膜区细胞膜内深1.1nm的门冬氨酸似为与配体的主要结合点。
N胆碱受体是由两个单体形成的二聚体。每个单体由5个亚基组成,包括两个α亚基(分子量为40000),一个β亚基(50000),一个γ基(57000)和一个δ亚基(64000)。每个亚基都有4个跨膜区段。5个亚基围绕成环状而形成离子通道(图5-8)。
二、传出神经按递质的分类
一般都根据所释放递质的不同,将传出神经分为胆碱能神经和去甲肾上腺素能神经两大类。
(一)胆碱能神经(cholinergicnerve)能合成Ach,兴奋时从末梢释放Ach。
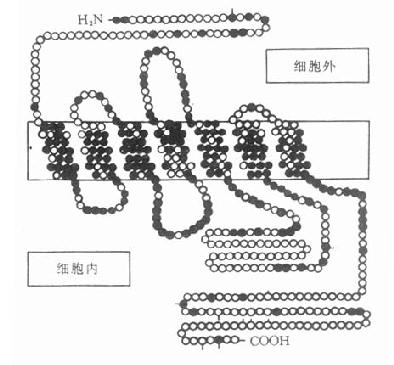
图5-7 人体β1-肾上腺受体
含477氨基酸,肽链跨膜7次,N端在细胞外,C端在
细胞内,胞内带短线的是丝氨酸,能被磷酸化。黑实心
圆是人体β1、β2受体所共有的氨基酸,在跨膜区较多。
(自TINS 11,321,1988)
1.全部交感神经和副交感神经的节前纤维;
2.运动神经;
3.全部副交感神经的节后纤维;
4.极少数交感神经节后纤维,如支配汗腺的分泌神经和骨骼肌的血管舒张神经。
(二)去甲肾上腺素能神经(noradrenergic nerve)能合成NA,兴奋时能释放NA。
几乎全部交感神经节后纤维都属此类。
传统认为一类神经元只释放一种递质,现知情况远较复杂。实际上许多神经元都贮有二或三种递质可供释放,例如颌下腺胆碱能神经元除有能促分泌的乙酰胆碱外,还随同释放血管活性肠肽(VIP),它能扩张血管,阿托品能抑制分泌,却不能拮抗血管扩张。大脑皮质神经元同此。肠壁神经元能共同释放VIP、强啡肽和甘丙肽。许多去甲肾上腺素能神经元也共同释放NA、ATP和神经肽Y。上述现象现称共同传递(cotransmission)。
三、传出神经系统效应产生的生化过程
神经递质或激动药受体结合后,触发一系列瀑布式的生化过程,通过一级一级地放大,最终导致效应,这一过程称为受体-效应偶联(receptor-effect coupling)。现介绍有关传出神经系统的两种受体反应偶联如下:
(一)受体与离子通道的偶联
神经递质或激动药与受体间相互作用可使受体操纵性离子通道(receptor-operatedclannel)开放,从而产生效应。例如β受体激动使钙离子通道开放,Ca2+进入细胞而产生各种生理效应。
有些受体含有离子通道,如上述N胆碱受体,与配体结合就能直接使通道开放。故称这种受体为门控通道型受体。
(二)受体与酶的偶联
传出神经系统的G蛋白偶联受体是通过腺苷酸环化酶(adenyl cyclase,AC)或磷脂酶C(phospholipase C, PLC)而产生效应的(图5-9)。
四、传出神经系统的生理功能
传出神经系统药物种类繁多,但它们药理作用的共性不外是影响传出神经系统的功能,或是拟似药,或是拮抗药。因此,如果熟悉两大类传出神经即去甲肾上腺素能神经和胆碱能神经的生理功能,再结合各药的特性,自易掌握每个药物的药理作用。
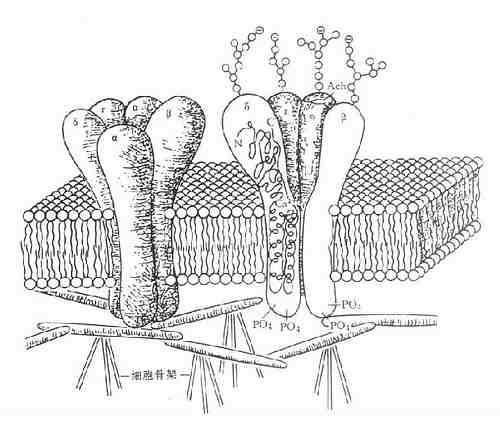
图5-8 N2烟碱受体
5个亚基各含约450个氨基酸,此5个肽链形成一个跨膜的环,在细胞内固定于细胞骨架上,每一肽链跨膜4次,N端和C端都位于胞外部(如δ亚单位剖面所示)。肽链在胞外被糖基化。在胞内被磷酸化,导致受体脱敏,2个α单位各有1个Ach结合位点,二者都结合1分子Ach后,钠通道开放,细胞除极兴奋。(引自F.Hucho)
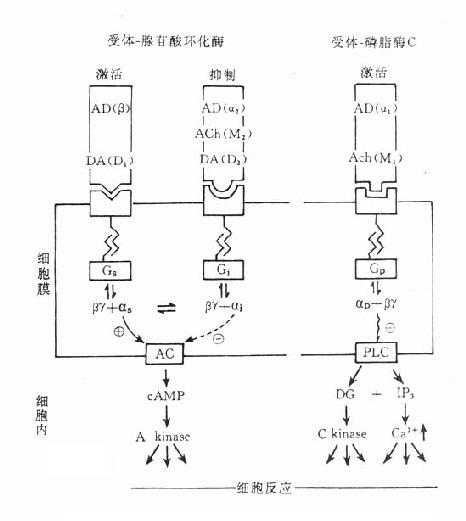
图5-9 受体-腺苷酸环化酶偶联和受体-磷脂酶偶联示意图
多数器官都接受上述两大类传出神经的双重支配。去甲肾上腺素能神经兴奋时(相当于递质去甲肾上腺素的作用),可见心脏兴奋、皮肤粘膜和内脏血管收缩、血压升高、支气管和胃肠道平滑肌抑制、瞳孔扩大等。这些功能变化,有利于机体适应环境的急聚变化。胆碱能神经兴奋时(相当于递质乙酰胆碱作用),节前与节后纤维的功能有所不同,当节后纤维兴奋时,基本上表现与上述相反的作用,有利于机体进行休整和积蓄能量。当节前纤维兴奋时,可引起神经节兴奋和肾上腺髓质分泌的增加,细节见表5-1。
表5-1 传出神经的受体-效应表*
| 效应器 | 肾上腺素能神经兴奋 | 胆碱能神经兴奋 |
| ||||
| 效 应 | 受 体 | 效应 | 受体 |
| |||
| 心脏 | 心肌 窦房结 传导系统 | 收缩力加强⑧ 心率加快 传导加快 | β1① | 收缩力减弱 心率减慢 传导减慢 | M M | ||
| 平 滑 肌 | 血 管 | 皮肤、粘膜 腹腔内脏② 骨骼肌 冠状动脉 | 收缩 收缩 舒张 收缩 舒张 舒张 | α α β2 α β2 β2 | 舒张④ 舒张(交感神经) | ||
| 支气管,气管 胃肠壁⑤ 膀胱逼尿肌 胃肠和膀胱括约肌 胆囊与胆道 | 舒张 舒张 舒张 收缩 舒张 | β2 α、β2 β2 α β2 | 收缩 收缩 收缩 舒张 收缩 | ||||
| 子宫 | 收缩⑥ 抑制 | α β2 | 不定 | ||||
| 眼 | 虹膜 睫状肌 | 瞳孔扩大肌收缩(扩瞳) 舒张(远视) | α β2 | 瞳孔括约肌收缩(缩瞳) 收缩(近视) | |||
| 腺 体 | 汗腺 唾液腺 胃肠道及呼吸道腺体 | 手心脚心分泌 分泌K+及H2O 分泌淀粉酶 | α′ α β2 | 全身分泌(交感神经) 分泌K+及H2O 分泌 | |||
| 代谢 | 肝脏糖代谢 骨骼肌糖代谢 脂肪代谢 | 肝糖原分解及异生 肌糖原分解 脂肪分解 | α、β2 β2 α、β2⑦ | ||||
| 植物神经节 肾上腺髓质 | 兴奋 分泌(交感神经节前纤维) | N1 | |||||
| 骨骼肌 | 收缩 | β2 | 收缩(运动神经) | N2 | |||
①心肌也具α和β2受体;人的心肌β1受体与β2受体之比约为80:20。
②肾脏和肠系膜血管尚有多巴胺受体,激动时血管舒张。
③虽也有α受体,但在整体动物,由于自身调节机制而表现为舒张。
④无生理意义。
⑤消化道神经丛副交感神经节前纤维末梢可能有α受体,激动时抑制乙酰胆碱的释放,产生抑制效应。
⑥雌激素占优势时,可致收缩。
⑦不同动物,受体亚型各异。
⑧黑体字表示占优势。
*许多组织都有不同型的受体共存,本表所列是在各组织中占多数的主要受体。
人体内的生理调节是在对立统一规律下进行的。在同一器官上,胆碱能神经和去甲肾上腺素能神经的作用大多是互相对抗的,但在中枢神经系统的调节下,它们的功能既是对立的,又是统一的。
近年来,在受体水平的研究中,也发现胆碱能神经和去甲肾上腺素能神经的功能并非截然分割,而是互相调节和互相制约的。例如有此去甲肾上腺素能神经和胆碱能神经突触前膜可能兼具抑制性的α受体和M受体,即受其本身所释放递质的反馈性调节,也受其生理拮抗性神经元所释放的递质的控制。
五、传出神经系统药物的基本作用
(一)直接作用于受体
许多传出神经系统药物能直接与胆碱受体或肾上腺素受体结合。结合后,如果产生与递质相似的作用,就称激动药。如果结合后不产生或较少产生拟似递质的作用,相反,却能妨碍递质与受体的结合,从而阻断了冲动的传递,产生与递质相反的作用,就称为阻断药(blocker);对激动药而言,可称拮抗药。
这类药物品种很多,也较常用。由于胆碱受体分为M和N两型,肾上腺素受体也有α和β两型。因此,选择性地作用于不同型受体的激动药和阻断药也具有相应的分类。
(二)影响递质
1.影响递质的生物合成直接影响递质生物合成的药物较少,且无临床应用价值,仅作药理学研究的工具药。
2.影响递质的转化如乙酰胆碱的灭活主要是被胆碱酯酶水解。因此,抗胆碱酯酶药就能妨碍乙酰胆碱的水解,提高其浓度,产生效应。
表5-2常用传出神经系统药物的分类
| 拟 似 药 | 拮 抗 药 |
| 1.M,N受体激动药(氨甲酰胆碱) 2.M受体激动药(毛果云香碱) 3.N受体激动药(烟碱) (二)抗胆碱酯酶药(新斯的明) (三)肾上腺素受体激动药 1.α受体激动药 (1)α1,α2受体激动药(去甲肾上腺素) (2)α1受体激动药(去氧肾上腺素) (3)α2受体激动药(可乐定) 2.α、β受体激动药(肾上腺素) 3.β受体激动药 (1)β1,β2受体激动药(异丙肾上腺素) (2)β1受体激动药(多巴酚丁胺) (3)β2受体激动药(沙丁胺醇) | (一)胆碱受体阻断药 1.M受体阻断药 (1)非选择性M受体阻断药(阿托品) (2)M1受体阻断药(哌仑西平) 2.N受体阻断药 (1)N1受体阻断药(六甲双铵) (2)N2受体阻断药(琥珀胆碱) (二)胆碱酯酶复活药(碘解磷定) (三)肾上腺素受体阻断药 1.α受体阻断药 (1)α1,α2受体阻断药 ①短效类(酚妥拉明) ②长效类(酚苄明) (2)α1受体阻断药(哌唑嗪) (3)α2受体阻断药(育亨宾) 2.β受体阻断药 1A类(普萘洛尔) 1B类(吲哚洛尔) 2A类(阿替洛尔) 2B类(艾司洛尔,esmolol,短效) 3类(拉贝洛尔) |
去甲肾上腺素作用的消失与乙酰胆碱不同,它主要靠突触前膜的摄取,因此现有的MAO抑制药或COMT抑制药并不能成为理想的外周拟肾上腺素药。
3.影响递质的转运和贮存药物可通过促进递质的释放而发挥递质样作用。例如麻黄碱促进去甲肾上腺素的释放、氨甲酰胆碱促进乙酰胆碱的释放而发挥作用,虽然它们同时尚有直接与受体结合的作用。
药物也可通过影响递质在神经末梢的贮存而发挥作用。例如利血平抑制神经末梢囊泡对去甲肾上腺素的摄取,使囊泡内去甲肾上腺素逐渐减少以至耗竭,从而表现为拮抗去甲肾上腺素能神经的作用。
六、传出神经系统药物的分类
传出神经系统药物可按其作用性质(激动受体或阻断受体)和对不同类型受体的选择性进行分类如表5-2。也可将影响胆碱酯酶的药单列一类。
(上海医科大学 杨藻宸)
-
《药理学》 中的相关章节:
……
第三节 药物消除动力学
第四章 影响药物效应的因素及合理用药原则
第一节 药物方面的因素
第二节 机体方面的因素
第三节 合理用药原则
第五章 传出神经系统药理概论(当前页)
第六章 胆碱受体激动药
第一节 M,N胆碱受体激动药
第二节 M胆碱受体激动药
第三节 N胆碱受体激动药
……