��Ѫ�����������������Ѫ��ϸ��HL-60����˿����ԭ�����ø�Ĺ�ϵ
�����ڶ���ҽ��ѧѧ��2000���21����6��
�����桡���̿�������Ƽ�������١�������
���� ��ժ��Ҫ��Ŀ��:̽������Ѫ��ϸ��ϵHL-60��Ѫ���������ӣ�LIF��������ǻ�(gp190)����һ�ǻ�(gp130)��ϸ����������˿����ԭ�����ø(MAPK)�Ĺ�ϵ���˽�LIF�����ڵ��ڰ�Ѫ��ϸ����ֳ�ͷֻ��е����塣����:�û������鼼����������ϸ�����������Թ�����Ƕ�������壨190/130��130/190�����ֱ���HL-60�������Ұ�������徺���Խ��LIF���������黯�͵�����ӡ������������ϸ�������γ�ͬԴ�Զ����壨190cyt-190cyt��130cyt-130cyt�����HL-60ϸ����MAPK p42/p44��Thr202/Tyr204���ữ״���������תȾpED190/130����LIF����10 min��HL-60ϸ��MAPK p42/p44 ��Thr202/Tyr204���ữ���ӣ�ϸ����ֳ���ԣ�����һǶ������������ǻ��γ�190cyt-190cytʱ hL-60ϸ��MAPK�����ữ���͡� ���ۣ� LIFR��gp130�ǻ���ϸ������������MAPK�ļ����Ѫ��ϸ��HL-60��ֳ�źŵĴ��ݡ�
�����ؼ��ʣ���Ѫ�����������ӣ����ߣ����壻HL-60����˿����ԭ������ø��
������Ѫ���������ӣ�LIF�������ư�Ѫ��ϸ��,��U937��HL-60��M1�ȵ���ֳ��1������ЧӦ�����ڽ�ϰ�ϸ��Ĥ�ϵ�LIF������ǻ�(gp190)����ʹ֮����һ�ǻ���gp130���γ���Դ�Զ�������2�����Ӷ�ʹϸ�����źŷ������ữ�������ữ��3�� ,�Ե���ϸ����DNA�ϳɺ�ϸ�������ٶȡ��о���һ��֤ʵLIF��������˿����ԭ�����ø(MAPK)��ϵ������4����Ȼ����LIF������һ�ǻ�Ӱ��MAPK�Ļ���?������Щ���Ӳ���?Ŀǰ�Բ���������о�������Ѫ��ϸ��HL-60Ϊ��ϸ�������ı���LIF����װ����ϸ��Ĥ�ϡ�ͨ�����ϸ����MAPK�ı����Լ����Ұ������ữˮƽ���ж�LIF�������ǻ���MAPK�Ĺ�ϵ������HL-60ϸ���ڵ���ϸ����ֳ�����á�
����1�����Ϻͷ���
����1.1��ϸ������HL-60ϸ�������п�Ժ�Ϻ�ϸ�����ṩ����ϸ�����ú�10%СţѪ���DMEM��Gibco��37�棬5% CO2 ������������
����1.2��LIF�������ǻ�ϸ�������Ļ�������pKCSR�� gp190 ��LIF ������ǻ�����pRclgo gp130��Moreau��ʿ�ṩ��Ϊģ�壬�ֱ���Sal��BamH���Sal��/Xho��Xba��ø�в�װ��pALTER���ϳ�����1:5��-CCATTCTCGTTTTCTAGAGCAAAGGAT-3�������2:5��-GTCTCGCTTTCTAGAGCAGAACAGCAC-3��;����Altered Sites ��ͻ��ϵͳ��Promega���ֱ�ʹ���ǻ��Ŀ�Ĥ����ϸ������֮�����Xba��ø��λ�㡣Ȼ����Xba���BamH��ȡ�¸���ϸ�������������л�����190/130��130/190�������Pst��,EcoR �� Pst ��,Sma �����ǻ�Ƕ�����pALTER������������pED��Moreau��ʿ�ṩ����
����1.3��LIF�������ǻ�Ƕ��������HL-60ϸ��תȾ����190/130��130/190Ƕ�������ķ�ת¼��������pED��pED190/130��pED 130/190�� ��֬����תȾ����Gibco-BRL��˾��LipofectamineTM�Ͳ������裩�ֱ�תȾ��HL-60��72 h�����LIF��5�� 100��250 ��g/ml��Ӧ10 min��4�����ģ��ռ�ϸ�����ڱ��ϡ�
����1.4�������黯���MAPK�����ữˮƽ�����䶳PBS��ϸ��ϴ���Σ����Ž���4%��ȩ�й̶�30 min������Ϳ���ڸ��и������������ز�Ƭ�ϣ�37���ҹ�����ǰ��Ϳ��ϸ�����ز�Ƭ���¹̶�20 min��Ȼ����PBSϴ3�Ρ��μ�������Ѫ����20 min����p42/p44 MAPK��Thr202/Tyr204�����ữ�ĵ���¡����(Pharmagene)37�淴Ӧ40 min ��AKP����ÿ���IgG(Promega)��ͬ����������ϸ���������������ڼ�������ø����Һ��pH9.5��5 min���� BCIP/NBTʹ֮��ɫ�������۲�ϸ����ɫ�ʵ���������ˮ��ϴ����ɡ�����������Ƭ��������ͼ�����ϵͳ�����գ�FR-988 smartScape 2000�����ࡢ��ǡ�������
����1.5��������ӡ��������MAPK�����ữ״�������䶳PBS��ϸ��ϴ���Σ�Ȼ��1��107��/mlϸ���ܶȼ�100��l TBS��pH 7.4�� 20 mmol/L Tris -HCl�� 150 mmol/L NaCl�� 1 mmol/L EDTA����1% triton X-100�� 200 ��mol/L PMSF��Sigma���� 2 ��mol/L Leupeptin�� 2 ��mol/L aprotinine����˳�Լ����ڱ��Ϸ�Ӧ20 min��4������ 12 000 r/min��20 min�� ������12% SDS-PAGE������Ӿ���ò�ʽתĤ�ǣ�TE52X Pharmacia biotech��������ת�Ƶ�PVDF��Whatman������p42/p44 MAPK���ữ����¡������Ĥ����ϣ�AKP��ǵĿ���IgG��һ����ϣ�BCIP/NBT��Ϊø������ɫ��
����1.6��ͳ��ѧ��������t���鴦������MAPK���ữ�仯�����ݡ�
����2���ᡡ��
����2.1��LIF����Ƕ����190/130��130/190���Ʊ�����Altered Sites ��ͻ��ϵͳ�������gp190��gp130�Ŀ�Ĥ����ϸ�����������������������øλ�㾭Xba��ø��֤ʵ�����ǻ���ϸ��������Ĥ����ϸ����������ֱ���ȫ�Զ�DNA�����ǣ�ABI pRISM 373A����������ȷ����
����2.2��LIF����Ƕ����190/130��130/190��תȾ����֬���彫pED190/130��pED130/190�ֱ�תȾ��HL-60ϸ��������תȾ pEGFP-N1 ��Clontech����3 d����ӫ�������۲�GFP�ı��60%��70%��ϸ��Ϊ���ԣ�������ӡ����֤ʵǶ����תȾ�ɹ���6����
����2.3�������黯���MAPK���ữ������100��250 ng/ml LIF��ϸ�������Ұ����LIF�����ǻ���Ƕ����130/190��190/130��϶��ֱ��γɶ����壨190cyt-190cyt��130cyt-130cyt�����۲����ϸ����MAPK�����ữˮƽ���������MAPK���ữˮƽ190/130��϶�����Ϊ�ߣ�P��0.01������130/190����ϵͣ�P��0.05��ͼ1����LIF���ò�ͬʱ�䣨0��30 min����Ұ����ϸ����MAPK���ữˮƽ�����½���190/130��5 min��MAPK���ữˮƽ������10 min �ﵽ���壻ͬʱ130/190��10 min��MAPK���ữˮƽ�½���15��20 min �½���Ϊ���ԣ�ͼ2����ʵ��֤����LIF ���յ�HL-60ϸ����MAPK���ữ�½�����gp130ϸ�������γɶ�����ʱ�ɼ���MAPK����һ�ǻ�gp190��֮�����ԣ�LIF������ЧӦ��Ҫ����gp190�鵼�ġ�
����2��4��������ӡ��������MAPK�Ļ��ԡ���LIF������תȾ�и�Ƕ�����ϸ����10��15 min������䶳������,�漴��Ӿ��תĤ��ӡ�ǡ����������190/130���Ⱦɫ�����϶������130/190���ͼ3����ʵ��֤����130ϸ�������γ�ͬԴ�Զ��������յ�MAPK�ļ��
����2.5��ϸ����������LIF������תȾǶ����190/130��ϸ���� 72 h�������190/130��ϸ�������϶���������87%��P��0.001������130/190���������������졣���������gp130ϸ������������ϸ����ֳ�źŵĴ��ݡ�

ͼ 1��p42/p44 MAPK���ữˮƽ
����fig 1��p42/p44 MAPK phosphorylation level
����P��0.05, ������P��0.01 vs wild type
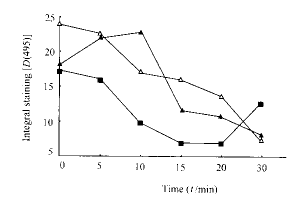
ͼ 2��LIF���ò�ͬʱ���ϸ����
����mAPK���ữˮƽ�ı仯
����fig 2��Various MAPK phosphorylation level in
����hL-60 induced by LIF at various time��: Wild type; ��: 190/130�� ��: 130/190
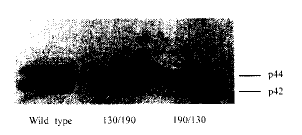
ͼ 3��������ӡ��������HL-60ϸ����
����mAPK p42/p44���ữ����
����fig 3��MAPK p42/p44 phosphorylation activity in
����hL-60 detected by Western blotting
����3���֡���
����LIF������ЧӦ��ͨ����ϰ�ϸ���ϵ�LIF������ǻ���gp190������ʹ֮��IL-6�źŴ����ӣ�gp130���γ���Դ�Զ����壬�Ӷ�����ϸ����һϵ���źŷ��ӵĻ��������LIF����Ϣ��ϸ��Ĥ��ϸ���˵Ĵ��ݣ�ʹϸ������һϵ�����������ı仯��3������Ȼ���屾�������ڵ���ø����ν֮�������Ұ��ἤø������壩�����ǣ��������γɽṹ�ı仯ʹJAK�ܹ���LIF�������ǻ��ϵ�Tyr���ữ����֮�γɡ��ٲ���ЧӦ������ sTATS�� MAPK���Լ�����ø����յ��͵��ڣ�����ʹϸ����ֳ�ͷֻ���
�����й�LIF������MAPK�Ĺ�ϵ������1994�걻Schiemann ����4��֤ʵ��������С��3T3-L1ϸ���ڵ�MAPK ����LIF ������ǻ���Ser-1044Ϊ��Ӧ������7�������������ֿ�չ������ضϷ���������������������ϸ�������γ�Ƕ��������ķ�������COS-7ϸ�����о�gp190��gp130ϸ��������ϸ����MAPK�Ĺ�ϵ,��Ϊgp190��gp130������MAPK�ļ���,�ɱ����ữλ��ֱ�ΪTyr118��Tyr115��8����
����HL-60ϸ��Ϊ��������ϸ����Ѫ��ϸ������������LIF���壬LIF�̼�������Ƹ�ϸ������ֳ��9�������о��������˽��Ѫ���ķ������ƣ���������Ѫ��ϵHL-60ϸ����Ϊ��ϸ�����о�תȾ190/130��130/190���ϸ������״̬�Լ�ϸ����MAPK�Ļ��Ա仯�������������LIF������ϸ��Ĥ�����壬��HL-60�����190/130��130/190�ֱ�ͬҰ���͵�gp190��gp130�����Խ��LIF���Ӷ��������ǻ�ϸ�������γ�ͬԴ�Զ�����ʱ��HL-60ϸ����Ӱ��,ǿ���ڰ�Ѫ��ϸ��ԭ����(����������ϸ�����źŷ���)�����ǰ����,����Ƕ�������ϸ�������ĸ�Ԥ���á����������LIF������HL-60ϸ��Ұ����LIF���壬ϸ����ֳ�ܵ����ƣ����δ�г�����LIF������תȾ190/130��HL-60ϸ����ϸ����ֳ�������ƣ�MAPK���ữ���ӣ���130/190��MAPK���ữ�½�����ˣ�������Ϊ��gp130��ϸ��������MAPK�ļ����йء�
����MAPK��ϸ���ڵ��źŴ��������ǽ������źţ�MEK��Raf-1��������Ĥ������ԭ������p42��p44 ��Thr202��Tyr204ͬʱ�����ữ����һê����(�����������Ĥ��ϣ������һ����10������������c-fos��ת¼����Ҫ�ٽ�ϸ����DNA�ϳ��Լ�ϸ���ķ��ѣ�ʹ��ϸ��������ֳ��11����130ϸ�������γ�ͬԴ�Զ�����ʱ��ǿ�ؼ���MAPK���������ź�;���б���LIF������ǻ���12���������ڵ���ϸ����ֳ�ͷֻ��з������á�
������������Ŀ��������Ȼ��ѧ����������Ŀ(39680012)
������������������棨1957�������У����壩�������ڣ�˶ʿ����ʦ�����ߵ�λ�������棨�ڶ���ҽ��ѧ����ҽѧ����֯��̥ѧ�����ң��Ϻ�200433)
�������̿����ڶ���ҽ��ѧ����ҽѧ����֯��̥ѧ�����ң��Ϻ�200433)
��������Ƽ���ڶ���ҽ��ѧ����ҽѧ����֯��̥ѧ�����ң��Ϻ�200433)
���������٣��ڶ���ҽ��ѧ����ҽѧ����֯��̥ѧ�����ң��Ϻ�200433)
���������꣨�ڶ���ҽ��ѧ����ҽѧ����֯��̥ѧ�����ң��Ϻ�200433��
���������
������1�ݡ�Metcalf d. The leukemia inhibitory factor��J��. Intern J Cell Cloning, 1991, 9(2):95-108.
������2�ݡ�Gearing DP, Thut CJ, VandaBos T, et al. Leukemia inhibitory factor receptor is structurally related to the IL-6 signal transducer gp130��J��. eMBO J, 1991,10(10):2839-2848.
������3�ݡ������棬������.��Ѫ�����������ķ��ӽṹ��ϸ������Ϣ������J��.����ҽѧ*��������ѧ�ֲᣬ1997��19(1):1-7.
������4�ݡ�Schiemann WP, Nathanson NM. Involvement of protein kinase C during activation of the mitogen-activated protein kinase cascade by leukemia inhibitory factor. Evidence for participation of multiple signaling pathways��J��. j Biol Chem,1994 ,269(9): 6376-6382.
������5�ݡ�Zhu K, Liu H, Liu XP, et al. Cloning and expression of leukemia inhibitory factor in CHO cell line and its inhibitory effect on the proliferation of U937��J��. J Exp Hematol, 1998,6(2):190-193.
������6�ݡ������棬����Ƽ��������,��.��Ѫ����������������gp130ϸ�������յ���Ѫ��ϸ��Meg-01��c-myc�ı����J��.�ڶ���ҽ��ѧѧ����2000,21(6):522-525.
������7�ݡ�Schiemann WP, Graves LM,Baumamann H, et al. Phosphoryaltion of the human leukemia inhibitory factor (LIF) receptor by mitogen-activated protein kinase and the regulation of LIF receptor function by heterologous receptor activation��J��. Proc Natl Acad Sci USA, 1995��92 (23) : 5361-5365.
������8�ݡ�Schiemann WP, Bartoe JL.Box 3-independent signaling mechanisms are involved in leukemia inhibitory factor receptor ��-and gp130-mediated stimualtion of mitogen-activated protein kinase��J��. J Biol Chem, 1997,272(26):16631-16636.
������9�ݡ�Maekawa T, Metcaff D. Colonial suppression of HL-60 and U937 cells by recombinant leukemia inhibitory factor in combination with GM-CSF or G-CSF��J��.Leukemia,1989,3(5):270-276.
������10�� Lenormand P, Brondello JM, Brunet A, et al. Growth factor-induced p42/p44 MAPK nuclear translocation and retention requires both MAPK activation and neosynthesis of nuclear anchoring proteins��J��. J Cell Biol, 1998,142(3): 625-633.
������11�� Porras A, Alvarez AM, Valaldares A, et al. p42/p44 mitogen-activated protein kinases activation is required for the insulin-like growth factor-��/insulin induced proliferation, but inhibits differentiation, in rat fetal brown adipocytes��J��. Mol Endocrinol, 1998, 12(6): 825-834.
������12�� Starr R, Novak U, Willson TA, et al .Distinct roles for leukemia inhibitory factor receptor ��-chain and gp130 in cell type-specific signal transduction��J��. J Biol Chem , 1997, 272(32): 19982-19986.