第二节 糖的分解代谢(catabolism of carbohydrate)
人体组织均能对糖进行分解代谢,主要的分解途径有四条:(1)无氧条件下进行的糖酵解途径;(2)有氧条件下进行的有氧氧化;(3)生成磷酸戊糖的磷酸戊糖通路;(4)生成葡萄糖醛酸的糖醛酸代谢。
一、糖酵解途径(glycolytic pathway)
糖酵解途径是指细胞在胞浆中分解葡萄糖生成丙酮酸(pyruvate)的过程,此过程中伴有少量ATP的生成。在缺氧条件下丙酮酸被还原为乳酸(lactate)称为糖酵解。有氧条件下丙酮酸可进一步氧化分解生成乙酰CoA进入三羧酸循环,生成CO2和H2O。
(一)葡萄糖的转运(transport of glucose)
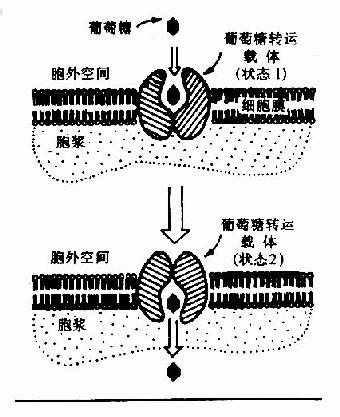
图4-1 葡萄糖通过转运载体转入细胞示意图GLUT代表葡萄糖转运载体
葡萄糖不能直接扩散进入细胞内,其通过两种方式转运入细胞:一种是在前一节提到的与Na+共转运方式,它是一个耗能逆浓度梯度转运,主要发生在小肠粘膜细胞、肾小管上皮细胞等部位;另一种方式是通过细胞膜上特定转运载体将葡萄糖转运入细胞内(图4-1),它是一个不耗能顺浓度梯度的转运过程。目前已知转运载体有5种,其具有组织特异性如转运载体-1(GLUT-1)主要存在于红细胞,而转运载体-4(GLUT-4)主要存在于脂肪组织和肌肉组织。
(二)糖酵解过程
糖酵解分为两个阶段共10个反应,每个分子葡萄糖经第一阶段共5个反应,消耗2个分子ATP为耗能过程,第二阶段5个反应生成4个分子ATP为释能过程。
1.第一阶段
(1)葡萄糖的磷酸化(phosphorylation of glucose)
进入细胞内的葡萄糖首先在第6位碳上被磷酸化生成6-磷酸葡萄糖(glucose6phophate,G-6-P),磷酸根由ATP供给,这一过程不仅活化了葡萄糖,有利于它进一步参与合成与分解代谢,同时还能使进入细胞的葡萄糖不再逸出细胞。催化此反应的酶是己糖激酶(hexokinase,HK)。己糖激酶催化的反应不可逆,反应需要消耗能量ATP,Mg2+是反应的激活剂,它能催化葡萄糖、甘露糖、氨基葡萄糖、果糖进行不可逆的磷酸化反应,生成相应的6-磷酸酯,6-磷酸葡萄糖是HK的反馈抑制物,此酶是糖氧化反应过程的限速酶(ratelimiting enzyme)或称关键酶(key enzyme)它有同工酶Ⅰ-Ⅳ型,Ⅰ、Ⅱ、Ⅲ型主要存在于肝外组织,其对葡萄糖Km值为10-5~10-6M。
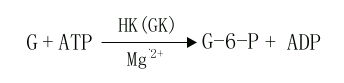
Ⅳ型主要存在于肝脏,特称葡萄糖激酶(glucokinase,GK),对葡萄糖的Km值1~10-2M,正常血糖浓度为5mmol/L,当血糖浓度升高时,GK活性增加,葡萄糖和胰岛素能诱导肝脏合成GK,GK能催化葡萄糖、甘露糖生成其6-磷酸酯,6-磷酸葡萄糖对此酶无抑制作用。
HK与GK两者区别见表4-1。
表4-1 己糖激酶(HK)和葡萄糖激酶(GK)的区别
| |
HK |
GK |
| 组织分布 |
绝大多数组织 |
肝脏和β细胞 |
| Km |
低 |
高 |
| 6-磷酸葡萄糖的抑制 |
有 |
无 |
(2)6-磷酸葡萄糖的异构反应(isomerization of glucose-6-phosphate)
这是由磷酸己糖异构酶(phosphohexose isomerase)催化6-磷酸葡萄糖(醛糖aldose sugar)转变为6-磷酸果糖(fructose-6-phosphate,F-6-P)的过程,此反应是可逆的。

(3)6-磷酸果糖的磷酸化(phosphorylation of fructose-6-phosphate)
此反应是6磷酸果糖第一位上的C进一步磷酸化生成1,6-二磷酸果糖,磷酸根由ATP供给,催化此反应的酶是磷酸果糖激酶1(phosphofructokinase l,PFK1)。
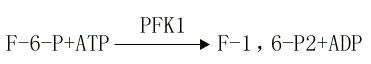
PFK1催化的反应是不可逆反应,它是糖的有氧氧化过程中最重要的限速酶,它也是变构酶,柠檬酸、ATP等是变构抑制剂,ADP、AMP、Pi、1,6-二磷酸果糖等是变构激活剂,胰岛素可诱导它的生成。
(4)1.6二磷酸果糖裂解反应(cleavage of fructose1,6 di/bis phosphate)
醛缩酶(aldolase)催化1.6-二磷酸果糖生成磷酸二羟丙酮和3-磷酸甘油醛,此反应是可逆的。

(5)磷酸二羟丙酮的异构反应(isomerization of dihydroxyacetonephosphate)
磷酸丙糖异构酶(triose phosphate isomerase)催化磷酸二羟丙酮转变为3-磷酸甘油醛,此反应也是可逆的。

到此1分子葡萄糖生成2分子3-磷酸甘油醛,通过两次磷酸化作用消耗2分子ATP。
2.第二阶段:
(6)3-磷酸甘油醛氧化反应(oxidation of glyceraldehyde-3-phosphate
此反应由3-磷酸甘油醛脱氢酶(glyceraldehyde 3-phosphatedehydrogenase)催化3-磷酸甘油醛氧化脱氢并磷酸化生成含有1个高能磷酸键的1,3-二磷酸甘油酸,本反应脱下的氢和电子转给脱氢酶的辅酶NAD+生成NADH+H+,磷酸根来自无机磷酸。

(7)1.3-二磷酸甘油酸的高能磷酸键转移反应
在磷酸甘油酸激酶(phosphaglycerate kinase,PGK)催化下,1.3-二磷酸甘油酸生成3-磷酸甘油酸,同时其C1上的高能磷酸根转移给ADP生成ATP,这种底物氧化过程中产生的能量直接将ADP磷酸化生成ATP的过程,称为底物水平磷酸化(substratelevel phosphorylation)。此激酶催化的反应是可逆的。
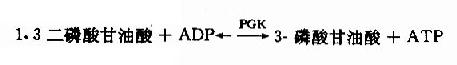
(8)3-磷酸甘油酸的变位反应
在磷酸甘油酸变位酶(phosphoglycerate mutase)催化下3-磷酸甘油酸C3-位上的磷酸基转变到C2位上生成2-磷酸甘油酸。此反应是可逆的。

(9)2-磷酸甘油酸的脱水反应
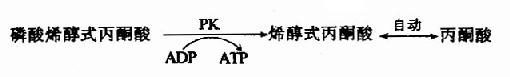
由烯醇化酶(enolase)催化,2-磷酸甘油酸脱水的同时,能量重新分配,生成含高能磷酸键的磷酸烯醇式丙酮酸(phosphoenolpyruvate PEP)。本反应也是可逆的。
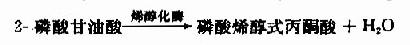
(10)磷酸烯醇式丙酮酸的磷酸转移
在丙酮酸激酶(pyruvate kinase,PK)催化下,磷酸烯醇式丙酮酸上的高能磷酸根转移至ADP生成ATP,这是又一次底物水平上的磷酸化过程。但此反应是不可逆的。
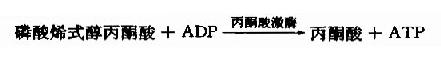
丙酮酸激酶是糖的有氧氧化过程中的限速酶,具有变构酶性质,ATP是变构抑制剂,ADP是变构激活剂,Mg2+或K+可激活丙酮酸激酶的活性,胰岛素可诱导PK的生成,烯醇式丙酮酸又可自动转变成丙酮酸。
总结糖的无氧酵解 在细胞液阶段的过程中,一个分子的葡萄糖或糖原中的一个葡萄糖单位,可氧化分解产生2个分子的丙酮酸,丙酮酸将进入线粒体继续氧化分解,此过程中产生的两对NADH+H+,由递氢体α-磷酸甘油(肌肉和神经组织细胞)或苹果酸(心肌或肝脏细胞)传递进入线粒体,再经线粒体内氧化呼吸链的传递,最后氢与氧结合生成水,在氢的传递过程释放能量,其中一部分以ATP形式贮存。
在整个细胞液阶段中的10或11步酶促反应中,在生理条件下有三步是不可逆的单向反应,催化这三步反应的酶活性较低,是整个糖的有氧氧化过程的关键酶,其活性大小,对糖的氧化分解速度起决定性作用,在此阶段经底物水平磷酸化产生四个分子ATP。
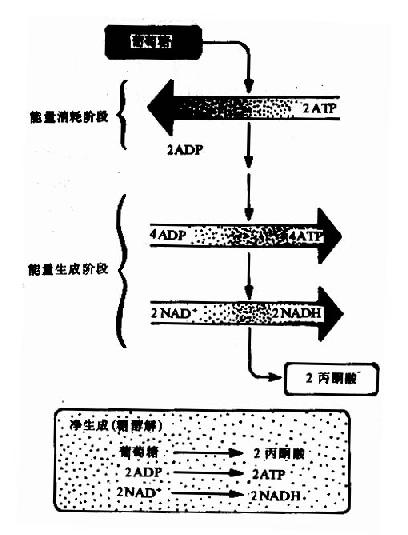
图4-2 葡萄糖分解的两个阶段
总而言之,经过糖酵解途径,一个分子葡萄糖可氧化分解产生2个分子丙酮酸。在此过程中,经底物水平磷酸化可产生4个分子ATP,如与第一阶段葡萄糖磷酸化和磷酸果糖的磷酸化消耗二分子ATP相互抵消,每分子葡萄糖降解至丙酮酸净产生2分子ATP,如从糖原开始,因开始阶段仅消耗1分子ATP,所以每个葡萄糖单位可净生成3分子ATP(图4-2)。葡萄糖+2Pi+2NAD++2ADP→2丙酮酸+2ATP+2NADH+2H++2H2O
(三)丙酮酸在无氧条件下生成乳酸
氧供应不足时从糖酵解途径生成的丙酮酸转变为乳酸。缺氧时葡萄糖分解为乳酸称为糖酵解(glycolysis),因它和酵母菌生醇发酵非常相似。丙酮酸转变成乳酸由乳酸脱氢酶(lactate dehydrogenase)催化丙酮酸乳酸脱氢酶乳酸在这个反应中丙酮酸起了氢接受体的作用。由3-磷酸甘油醛脱氢酶反应生成的NADH+H+,缺氧时不能经电子传递链氧化。正是通过将丙酮酸还原成乳酸,使NADH转变成NAD+,糖酵解才能继续进行。
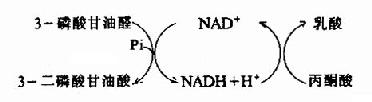 乳酸脱氢酶是由M和H二种亚基构成的四聚体,组成5种同工酶。这些同工酶在组织中分布不同,对丙酮酸的KM也有较大差异。H4主要分布在心肌。它的酶动力学参数表明H4有利于催化乳酸氧化成丙酮酸。所以心肌进行有氧氧化而且能利用乳酸作为燃料。骨骼肌中为M4型。它对反应方面无倾向性,但肌细胞内底物的浓度有利于生成乳酸。
乳酸脱氢酶是由M和H二种亚基构成的四聚体,组成5种同工酶。这些同工酶在组织中分布不同,对丙酮酸的KM也有较大差异。H4主要分布在心肌。它的酶动力学参数表明H4有利于催化乳酸氧化成丙酮酸。所以心肌进行有氧氧化而且能利用乳酸作为燃料。骨骼肌中为M4型。它对反应方面无倾向性,但肌细胞内底物的浓度有利于生成乳酸。
(四)糖酵解及其生理意义
糖酵解是生物界普遍存在的供能途径,但其释放的能量不多,而且在一般生理情况下,大多数组织有足够的氧以供有氧氧化之需,很少进行糖酵解,因此这一代谢途径供能意义不大,但少数组织,如视网膜、睾丸、肾髓质和红细胞等组织细胞,即使在有氧条件下,仍需从糖酵解获得能量。
在某些情况下,糖酵解有特殊的生理意义。例如剧烈运动时,能量需求增加,糖分解加速,此时即使呼吸和循环加快以增加氧的供应量,仍不能满足体内糖完全氧化所需要的能量,这时肌肉处于相对缺氧状态,必须通过糖酵解过程,以补充所需的能量。在剧烈运动后,可见血中乳酸浓度成倍地升高,这是糖酵解加强的结果。又如人们从平原地区进入高原的初期,由于缺氧,组织细胞也往往通过增强糖酵解获得能量。
在某些病理情况下,如严重贫血、大量失血、呼吸障碍、肿瘤组织等,组织细胞也需通过糖酵解来获取能量。倘若糖酵解过度,可因乳酸产生过多,而导致酸中毒。
(五)糖酵解的调节
正常生理条件下,人体内的各种代谢受到严格而精确的调节,以满足机体的需要,保持内环境的稳定。这种控制主要是通过调节酶的活性来实现的。在一个代谢过程中往往催化不可逆反应的酶限制代谢反应速度,这种酶称为限速酶。糖酵解途径中主要限速酶是己糖激酶(HK),磷酸果糖激酶-1(PFK-1)和丙酮酸激酶(PK)。
1.激素的调节
胰岛素能诱导体内葡萄糖激酶、磷酸果糖激酶、丙酮酸激酶的合成,因而促进这些酶的活性,一般来说,这种促进作用比对限速酶的变构或修饰调节慢,但作用比较持久。
2.代谢物对限速酶的变构调节
上述三个限速酶中,起决定作用的是催化效率最低的酶PFK-1。其分子是一个四聚体形式,不仅具有对反应底物6-磷酸果糖和ATP的结合部位,而且尚有几个与别位激活剂和抑制剂结合的部位,6-磷酸果糖、1,6二磷酸果糖、ADP和AMP是其激活剂,而ATP、柠檬酸等是其抑制剂,ATP既可作为反应底物又可作为抑制剂,其原因在于:此酶一个是与作为底物的ATP结合位点,另一个是与作为抑制剂的ATP结合位点,两个位点对ATP的亲和力不同,与底物的位点亲和力高,抑制剂作用的位点亲和力低。对ATP有两种结合位点,这样,当细胞内ATP不足时,ATP主要作为反应底物,保证酶促反应进行,而当细胞内ATP增多时,ATP作为抑制剂,降低了酶对6-磷酸果糖的亲和力。
它在体内也是由6-磷酸果糖磷酸化而成,但磷酸化是在C2位而不是C4位,参与的酶也是另一个激酶,磷酸果糖激酶-2(PFK-2)。
2,6-二磷酸果糖可被二磷酸果糖磷酸酶-2去磷酸而生成6-磷酸果糖,失去其调节作用。2,6-二磷酸果糖的作用在于增强磷酸果糖激酶-1对6-磷酸果糖的亲和力和取消ATP的抑制作用(图4-3)。
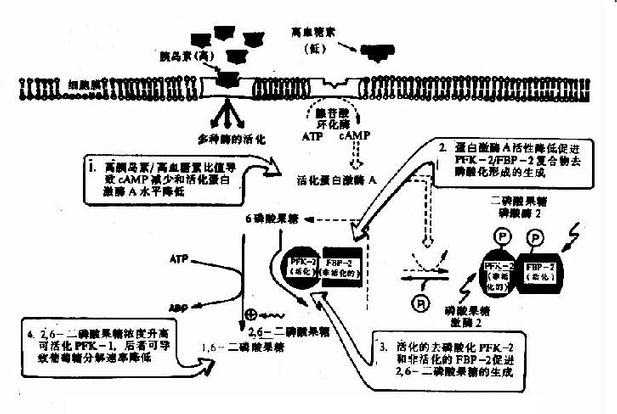
图4-3 胰岛素浓度升高对肝细胞内2,6-二磷酸果糖浓度的影响
临床上丙酮酸激酶异常,可导致葡萄糖酵解障碍,红细胞破坏出现溶血性贫血。
二、糖的有氧氧化
葡萄糖在有氧条件下,氧化分解生成二氧化碳和水的过程称为糖的有氧氧化(aerobicoxidation)。有氧氧化是糖分解代谢的主要方式,大多数组织中的葡萄糖均进行有氧氧化分解供给机体能量。
(一)有氧氧化过程
糖的有氧氧化分两个阶段进行。第一阶段是由葡萄糖生成的丙酮酸,在细胞液中进行。第二阶段是上述过程中产生的NADH+H+和丙酮酸在有氧状态下,进入线粒体中,丙酮酸氧化脱羧生成乙酰CoA进入三羧酸循环,进而氧化生成CO2和H2O,同时NADH+H+等可经呼吸链传递,伴随氧化磷酸化过程生成H2O和ATP,下面主要将讨论有氧氧化在线粒体中进行的第二阶段代谢。
1.丙酮酸的氧化脱羧
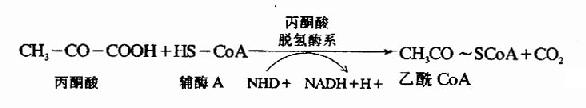
催化氧化脱羧的酶是丙酮酸脱氢酶系(pyruvate dehydrogenase system),此多酶复合体括丙酮酸脱羧酶,辅酶是TPP,二氢硫辛酸乙酰转移酶,辅酶是二氢硫辛酸和辅酶A,还有二氢硫辛酸脱氢酶,辅酶是FAD及存在于线粒体基质液中的NAD+,多酶复合体形成了紧密相连的连锁反应机构,提高了催化效率。
从丙酮酸到乙酰CoA是糖有氧氧化中关键的不可逆反应,催化这个反应的丙酮酸脱氢酶系受到很多因素的影响,反应中的产物,乙酰CoA和NADH++H+可以分别抑制酶系中的二氢硫辛酸乙酰转移酶和二氢硫辛酸脱氢酶的活性,丙酮酸脱羧酶(pyruvate decarboxylase,PDC)活性受ADP和胰岛素的激活,受ATP的抑制。
丙酮酸脱氢反应的重要特征是丙酮酸氧化释放的自由能贮存在乙酰CoA中的高能硫酯键中,并生成NADH+H+(图4-4)。
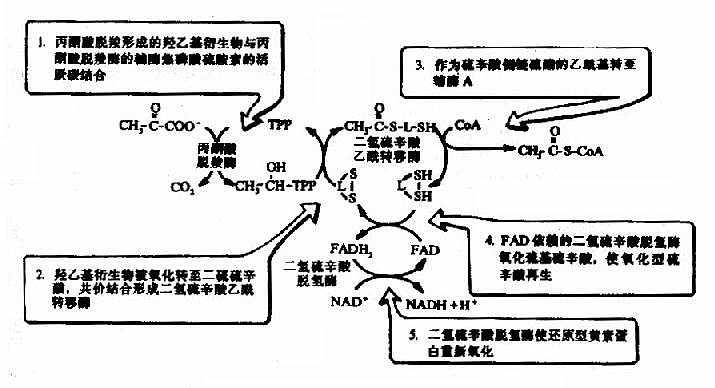
图4-4 丙酮酸脱氢酶复合物的作用机制

2.三羧酸循环(tricarboxylic acid cycle)
乙酰CoA进入由一连串反应构成的循环体系,被氧化生成H2O和CO2。由于这个循环反应开始于乙酰CoA与草酰乙酸(oxaloacetate)缩合生成的含有三个羧基的柠檬酸,因此称之为三羧酸循环或柠檬酸循环(citric acid cycle)。其详细过程如下:
(1)乙酰CoA进入三羧酸循环
乙酰CoA具有硫酯键,乙酰基有足够能量与草酰乙酸的羧基进行醛醇型缩合。首先从CH3CO基上除去一个H+,生成的阴离子对草酰乙酸的羰基碳进行亲核攻击,生成柠檬酰CoA中间体,然后高能硫酯键水解放出游离的柠檬酸,使反应不可逆地向右进行。该反应由柠檬酸合成酶(citrate synthetase)催化,是很强的放能反应。

由草酰乙酸和乙酰CoA合成柠檬酸是三羧酸循环的重要调节点,柠檬酸合成酶是一个变构酶,ATP是柠檬酸合成酶的变构抑制剂,此外,α-酮戊二酸、NADH能变构抑制其活性,长链脂酰CoA也可抑制它的活性,AMP可对抗ATP的抑制而起激活作用。
(2)异柠檬酸形成
柠檬酸的叔醇基不易氧化,转变成异柠檬酸而使叔醇变成仲醇,就易于氧化,此反应由顺乌头酸酶催化,为一可逆反应。

(3)第一次氧化脱酸
在异柠檬酸脱氢酶作用下,异柠檬酸的仲醇氧化成羰基,生成草酰琥珀酸(oxalosuccinate)的中间产物,后者在同一酶表面,快速脱羧生成α-酮戊二酸(αketoglutarate)、NADH和CO2,此反应为β-氧化脱羧,此酶需要Mn2+作为激活剂。

此反应是不可逆的,是三羧酸循环中的限速步骤,ADP是异柠檬酸脱氢酶的激活剂,而ATP,NADH是此酶的抑制剂。
(4)第二次氧化脱羧
在α-酮戊二酸脱氢酶系作用下,α-酮戊二酸氧化脱羧生成琥珀酰CoA、NADH+H+和CO2,反应过程完全类似于丙酮酸脱氢酶系催化的氧化脱羧,属于α氧化脱羧,氧化产生的能量中一部分储存于琥珀酰CoA的高能硫酯键中。
α-酮戊二酸脱氢酶系也由三个酶(α-酮戊二酸脱羧酶、硫辛酸琥珀酰基转移酶、二氢硫辛酸脱氢酶)和五个辅酶(TPP、硫辛酸、HSCoA、NAD+、FAD)组成。

此反应也是不可逆的。α-酮戊二酸脱氢酶复合体受ATP、GTP、NAPH和琥珀酰CoA抑制,但其不受磷酸化/去磷酸化的调控。
(5)底物磷酸化生成ATP
在琥珀酸硫激酶(succinate thiokinase)的作用下,琥珀酰CoA的硫酯键水解,释放的自由能用于合成GTP,在细菌和高等生物可直接生成ATP,在哺乳动物中,先生成GTP,再生成ATP,此时,琥珀酰CoA生成琥珀酸和辅酶A。

(6)琥珀酸脱氢
琥珀酸脱氢酶(succinate dehydrogenase)催化琥珀酸氧化成为延胡索酸。该酶结合在线粒体内膜上,而其他三羧酸循环的酶则都是存在线粒体基质中的,这酶含有铁硫中心和共价结合的FAD,来自琥珀酸的电子通过FAD和铁硫中心,然后进入电子传递链到O2,丙二酸是琥珀酸的类似物,是琥珀酸脱氢酶强有力的竞争性抑制物,所以可以阻断三羧酸循环。

(7)延胡索酸的水化
延胡索酸酶仅对延胡索酸的反式双键起作用,而对顺丁烯二酸(马来酸)则无催化作用,因而是高度立体特异性的。
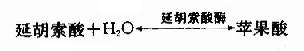
(8)草酰乙酸再生
在苹果酸脱氢酶(malic dehydrogenase)作用下,苹果酸仲醇基脱氢氧化成羰基,生成草酰乙酸(oxalocetate),NAD+是脱氢酶的辅酶,接受氢成为NADH+H+(图4-5)。
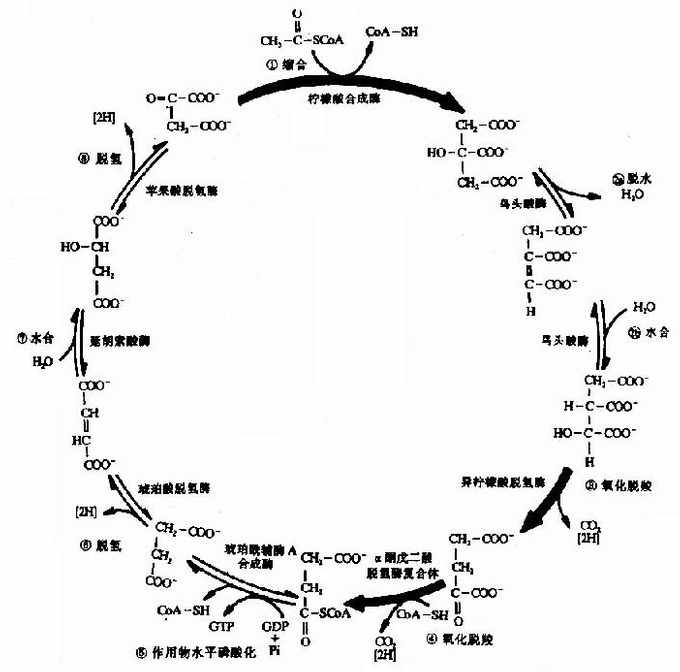
图4-5 三羧酸循环
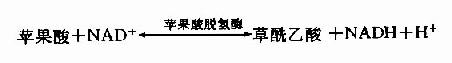
三羰酸循环总结:
乙酰CoA+3NADH++FAD+GDP+Pi+2H2O—→
2CO2+3NADH+FADH2+GTP+3H+ +CoASH
①CO2的生成,循环中有两次脱羧基反应(反应3和反应4)两次都同时有脱氢作用,但作用的机理不同,由异柠檬酸脱氢酶所催化的β氧化脱羧,辅酶是NAD+,它们先使底物脱氢生成草酰琥珀酸,然后在Mn2+或Mg2+的协同下,脱去羧基,生成α-酮戊二酸。
α-酮戊二酸脱氢酶系所催化的α氧化脱羧反应和前述丙酮酸脱氢酶系所催经的反应基本相同。
应当指出,通过脱羧作用生成CO2,是机体内产生CO2的普遍规律,由此可见,机体CO2的生成与体外燃烧生成CO2的过程截然不同。
②三羧酸循环的四次脱氢,其中三对氢原子以NAD+为受氢体,一对以FAD为受氢体,分别还原生成NADH+H+和FADH2。它们又经线粒体内递氢体系传递,最终与氧结合生成水,在此过程中释放出来的能量使ADP和Pi结合生成ATP,凡NADH+H+参与的递氢体系,每2H氧化成一分子H2O,生成3分子ATP,而FADH2参与的递氢体系则生成2分子ATP,再加上三羧酸循环中有一次底物磷酸化产生一分子ATP,那么,一分子CH2COSCoA参与三羧酸循环,直至循环终末共生成12分子ATP。
③乙酰CoA中乙酰基的碳原子,乙酰CoA进入循环,与四碳受体分子草酰乙酸缩合,生成六碳的柠檬酸,在三羧酸循环中有二次脱羧生成2分子CO2,与进入循环的二碳乙酰基的碳原子数相等,但是,以CO2方式失去的碳并非来自乙酰基的两个碳原子,而是来自草酰乙酸。
④三羧酸循环的中间产物,从理论上讲,可以循环不消耗,但是由于循环中的某些组成成分还可参与合成其他物质,而其他物质也可不断通过多种途径而生成中间产物,所以说三羧酸循环组成成分处于不断更新之中。
例如 草楚酰乙酸——→天门冬氨酸
α-酮戊二酸——→谷氨酸
草酰乙酸——→丙酮酸——→丙氨酸
其中丙酮酸羧化酶催化的生成草酰乙酸的反应最为重要。
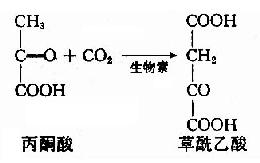
因为草酰乙酸的含量多少,直接影响循环的速度,因此不断补充草酰乙酸是使三羧酸循环得以顺利进行的关键。
三羧酸循环中生成的苹果酸和草酰乙酸也可以脱羧生成丙酮酸,再参与合成许多其他物质或进一步氧化(图4-6)。

(二)糖有氧氧化的生理意义
1.三羧酸循环是机体获取能量的主要方式。1个分子葡萄糖经无氧酵解仅净生成2个分子ATP,而有氧氧化可净生成38个ATP(如表4?),其中三羧酸循环生成24个ATP,在一般生理条件下,许多组织细胞皆从糖的有氧氧化获得能量。糖的有氧氧化不但释能效率高,而且逐步释能,并逐步储存于ATP分子中,因此能的利用率也很高。
2.三羧酸循环是糖,脂肪和蛋白质三种主要有机物在体内彻底氧化的共同代谢途径,三羧酸循环的起始物乙酰辅酶A,不但是糖氧化分解产物,它也可来自脂肪的甘油、脂肪酸和来自蛋白质的某些氨基酸代谢,因此三羧酸循环实际上是三种主要有机物在体内氧化供能的共同通路,估计人体内2/3的有机物是通过三羧酸循环而被分解的。
3.三羧酸循环是体内三种主要有机物互变的联结机构,因糖和甘油在体内代谢可生成α-酮戊二酸及草酰乙酸等三羧酸循环的中间产物,这些中间产物可以转变成为某些氨基酸;而有些氨基酸又可通过不同途径变成α-酮戊二酸和草酰乙酸,再经糖异生的途径生成糖或转变成甘油,因此三羧酸循环不仅是三种主要的有机物分解代谢的最终共同途径,而且也是它们互变的联络机构。
(三)糖有氧氧化的调节
如上所述糖有氧氧化分为两个阶段,第一阶段糖酵解途径的调节在糖酵解部分已探讨过,下面主要讨论第二阶段丙酸酸氧化脱羧生成乙酰CoA并进入三羧酸循环的一系列反应的调节。丙酮酸脱氢酶复合体、柠檬酸合成酶、异柠檬酸脱氢酶和α-酮戊二酸脱氢酶复合体是这一过程的限速酶。
丙酮酸脱氢酶复合体受别位调控也受化学修饰调控,该酶复合体受它的催化产物ATP、乙酰CoA和NADH有力的抑制,这种别位抑制可被长链脂肪酸所增强,当进入三羧酸循环的乙酰CoA减少,而AMP、辅酶A和NAD+堆积,酶复合体就被别位激活,除上述别位调节,在脊椎动物还有第二层次的调节,即酶蛋白的化学修饰,PDH含有两个亚基,其中一个亚基上特定的一个丝氨酸残基经磷酸化后,酶活性就受抑制,脱磷酸化活性就恢复,磷酸化-脱磷酸化作用是由特异的磷酸激酶和磷酸蛋白磷酸酶分别催化的,它们实际上也是丙酮酸酶复合体的组成,即前已述及的调节蛋白,激酶受ATP别位激活,当ATP高时,PDH就磷酸化而被激活,当ATP浓度下降,激酶活性也降低,而磷酸酶除去PDH上磷酸,PDH又被激活了。
对三羧酸循环中柠檬酸合成酶、异柠檬酸脱氢酶和α-酮戊二酸脱氢酶的调节,主要通过产物的反馈抑制来实现的,而三羧酸循环是机体产能的主要方式。因此ATP/ADP与NADH/NAD+两者的比值是其主要调节物。ATP/ADP比值升高,抑制柠檬酸合成酶和异柠檬酶脱氢酶活性,反之ATP/ADP比值下降可激活上述两个酶。NADH/NAD+比值升高抑制柠檬酸合成酶和α-酮戊二酸脱氢酶活性,除上述ATP/ADP与NADH/NAD+之外其它一些代谢产物对酶的活性也有影响,如柠檬酸抑制柠檬酸合成酶活性,而琥珀酰CoA抑制α-酮戊二酸脱氢酶活性。总之,组织中代谢产物决定循环反应的速度,以便调节机体ATP和NADH浓度,保证机体能量供给。(图4-7)。
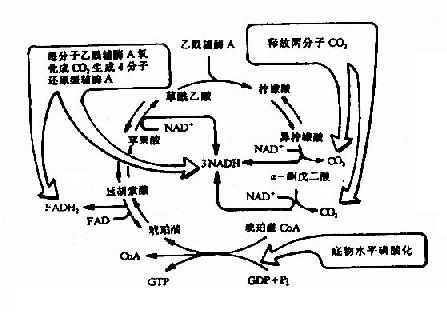
图4-6 三羧酸循环中还原型辅酶和CO2的生成
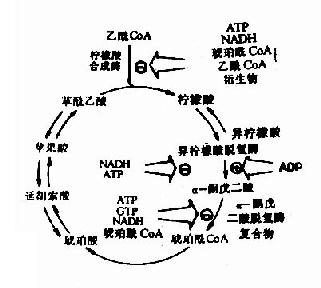
图4-7 三羧酸循环的抑制剂和激活剂
(四)有氧氧化和糖酵解的相互调节
Pasteur在研究酵母发酵时,发现在供氧充足的条件下,细胞内糖酵解作用受到抑制。葡萄糖消耗和乳酸生成减少,这种有氧氧化对糖酵解的抑制作用称为巴士德效应(Pasteureffect)。
产生巴士德效应主要是由于在供氧充足的条件下,细胞内ATP/ADP比值升高,抑制了PK和PFK,使6-磷酸果糖和6-磷酸葡萄糖含量增加,后者反馈抑制已糖激权衡利弊(HK),使葡萄糖利用减少,呈现有氧氧化对糖酵解的抑制作用。
Crabtree效应与巴士德效应相反,在癌细胞发现给予葡萄糖时不论供氧充足与否都呈现很强的酵解反应,而糖的有氧氧化受抑制,称为Crabtree效应或反巴士德效应。这种现象较普遍地存在于癌细胞中,此外也存在于一些正常组织细胞如视网膜、睾丸、颗粒白细胞等。
一般认为,具Crabtree效应的细胞,其酵解酶系(如PK、PFK、HK+)活性强,而线粒体内氧化酶系如细胞色素氧化酶活性则较低,它们在争夺ADF、Pi及ADH+H+方面线粒体必然处于劣势,因而缺乏进行氧化磷酸化的底物,即使在供氧充足的情况下,其有氧氧化生成ATP的能力仍低于正常细胞,呈现Crabtree效应。
三、磷酸戊糖途径
磷酸戊糖途径(pentose phosphate pathway)又称已糖单磷酸旁路(hexose monophosphate shut HMS)或磷酸葡萄糖旁路(phosphogluconate shut)。此途径由6-磷酸葡萄糖开始生成具有重要生理功能的NADPH和5-磷酸核糖。全过程中无ATP生成,因此此过程不是机体产能的方式。其主要发生在肝脏、脂肪组织、哺乳期的乳腺、肾上腺皮质、性腺、骨髓和红细胞等。
(一)反应过程
磷酸戊糖途径在细胞液中进行,全过程分为不可逆的氧化阶段和可逆的非氧化阶段。在氧化阶段,3个分子6-磷酸葡萄糖在6-磷酸葡萄糖脱氢酶和6-磷酸葡萄糖酸脱氢酶等催化下经氧化脱羧生成6个分子NADPH+H+,3个分子CO2和3个分子5-磷酸核酮糖;在非氧化阶段,5-磷酸核酮糖在转酮基酶(TPP为辅酶)和转硫基酶催化下使部分碳链进行相互转换,经三碳、四碳、七碳和磷酸酯等,最终生成2分子6-磷酸果糖和1分子3-磷酸甘油,它们可转变为6-磷酸葡萄糖继续进行磷酸戊糖途径,也可以进入糖有氧氧化或糖酵解途径(图4-8)。此反应途径中的限速酶是6-磷酸葡萄糖脱氢酶,此酶活性受NADPH浓度影响,NADPH浓度升高抑制酶的活性,因此磷酸戊糖途径主要受体内NADPH的需求量调节。
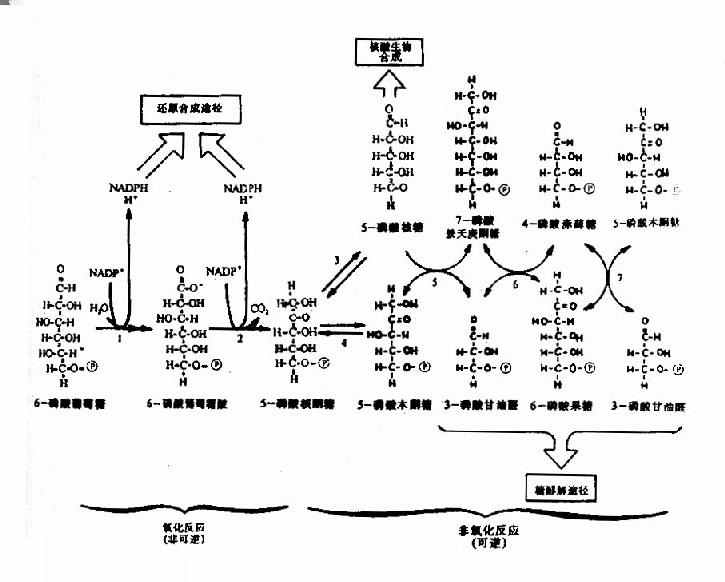
图4-8 已糖磷酸支路反应
(二)生理意义
1.5-磷酸核糖的生成,此途径是葡萄糖在体内生成5-磷酸核糖的唯一途径,故命名为磷酸戊糖通路,体内需要的5-磷酸核糖可通过磷酸戊糖通路的氧化阶段不可逆反应过程生成,也可经非氧化阶段的可逆反应过程生成,而在人体内主要由氧化阶段生成,5-磷酸核糖是合成核苷酸辅酶及核酸的主要原料,故损伤后修复、再生的组织(如梗塞的心肌、部分切除后的肝脏),此代谢途径都比较活跃。
2.NADPH+H+与NADH不同,它携带的氢不是通过呼吸链氧化磷酸化生成ATP,而是作为供氢体参与许多代谢反应,具有多种不同的生理意义。
(1)作为供氢体,参与体内多种生物合成反应,例如脂肪酸、胆固醇和类固醇激素的生物合成,都需要大量的NADPH+H+,因此磷酸戊糖通路在合成脂肪及固醇类化合物的肝、肾上腺、性腺等组织中特别旺盛。
(2)NADPH+H+是谷胱甘肽还原酶的辅酶,对维持还原型谷胱甘肽(GSH)的正常含量,有很重要的作用,GSH能保护某些蛋白质中的巯基,如红细胞膜和血红蛋白上的SH基,因此缺乏6-磷酸葡萄糖脱氢酶的人,因NADPH+H+缺乏,GSH含量过低,红细胞易于破坏而发生溶血性贫血。
(3)NADPH+H+参与肝脏生物转化反应,肝细胞内质网含有以NADPH+H+为供氢体的加单氧酶体系,参与激素、药物、毒物的生物转化过程。
(4)NADPH+H+参与体内嗜中性粒细胞和巨噬细胞产生离子态氧的反应,因而有杀菌作用。
四、糖醛酸代谢
糖醛酸代谢(uronic acid metabolism)主要在肝脏和红细胞中进行,它由尿嘧啶核苷二磷酸葡萄糖(UDPG)上联糖原合成途径,经过一系列反应后生成磷酸戊糖而进入磷酸戊糖通路,从而构成糖分解代谢的另一条通路。
1-磷酸葡萄糖和尿嘧啶核苷三磷酸(UTP)在尿二磷葡萄糖焦磷酸化酶(UDPG焦磷酸化酶)催化下生成尿二磷葡萄糖(UDPG),UDPG经尿二磷葡萄糖脱氢酶的作用进一步氧化脱氢生成尿二磷葡萄糖醛酸,脱氢酶的辅酶是NAD+,尿二磷葡萄糖醛酸(UDPGA)脱去尿二磷生成葡萄糖醛酸(glucuronic acid)。葡萄糖醛酸在一系列酶作用下,经NADPH+H+供氢和NAD+受氢的二次还原和氧化的过程,生成5-磷酸木酮糖进入磷酸戊糖通路(图4-9)。
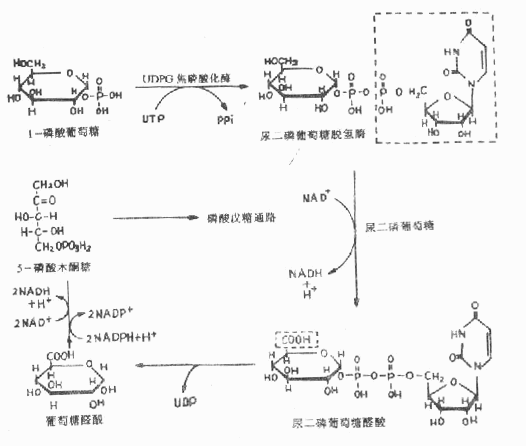
图4-9 糖醛酸代谢
糖醛酸代谢的主要生理功能在于代谢过程中生成了尿二磷葡萄糖醛酸,它是体内重要的解毒物质之一(详见肝脏生化章),同时又是合成粘多糖的原料(见结缔组织章)。此代谢过程要消耗NADPH+H+(同时生成NADH+H+),而磷酸戊糖通路又生成NADPH+H+,因此两者关系密切,当磷酸戊糖通路发生障碍时,必然会影响糖醛酸代谢的顺利进行。
五、其它单糖的酵解
人体可吸收利用的单糖除了葡萄糖以外,还有果糖和半乳糖等单糖,它们均可以通过转变过程,最终进入糖酵解途径。
果糖主要由蔗糖分解生成,存在于水果、蔬菜、蜂蜜中。它可被己糖激酶磷酸化,生成6-酸果糖而进入糖酵解途径。

这是体内各种组织均可进行的果糖磷酸化,但是由于己糖激酶对果糖的亲和力远远低于对葡萄糖的亲和力,因此在正常,以葡萄糖为主的食物代谢时,组织对果糖的磷酸化效率是很低的。只有在高果糖食物时,此代谢才进行。除上述反应外,果糖还有另一个磷酸化反应由果糖激酶(fructokinase)催化,此酶主要存在于肝、肾、小肠细胞中,本反应催化果糖在C1位上磷酸化生成1磷酸果糖。
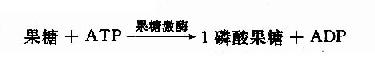
1-磷酸果糖可进一步由醛缩酶B催化裂解形成磷酸二羟丙酮和甘油醛,磷酸二羟丙酮可进入糖酵解途径。甘油醛可进一步被丙糖激酶催化生成3-磷酸甘油醛。
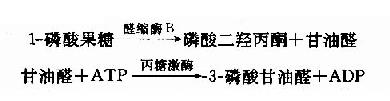
通过上述反应果糖可转变为糖酵解过程中的中间产物而进入糖酵解代谢途径。在上述反应中,限制果糖代谢的主要酶是磷酸酶B,此酶催化效率低,过量果糖可引起1磷酸果糖堆积而导致肝脏损害,出现低血糖、呕吐、黄疸、出血等以及高尿酸症和痛风。半乳糖来自牛乳中乳糖的水解,可由半乳糖激酶催化生成1磷酸半乳糖。
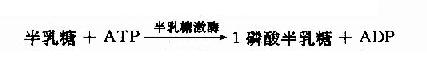
1-磷酸半乳糖可在1-磷酸半乳糖尿苷酰转移酶催化下与尿嘧啶核苷二磷酸葡萄糖(UDPG)作用,生成1磷酸葡萄糖和尿嘧啶核苷二磷酸半乳糖(UDp Gal)。
UDPG+1磷酸半乳糖←→1-磷酸葡萄糖+UDPGal
1-磷酸葡萄糖可在葡萄糖变位酶催化下转变为6-磷酸葡萄糖
1-磷酸葡萄糖←→6-磷酸葡萄糖
而UDPGal可在UDPG差向异构酶催化下转变为UDPG
UDPGal←→UDPG
这样1分子半乳糖可转变为1分子6-磷酸葡萄糖而进入酵解反应,而DPG无净消耗。1-磷酸
半乳糖尿苷酰转移酶缺乏可引起类似果糖代谢障碍出现的临床症状,而且影响更广泛可出现神经系统障碍,反应迟钝,尤其象脑、红细胞等特殊组织。