第五节 神经系统对内脏活动的调节
一、自主神经系统
自主神经系统是指调节内脏功能的神经装置,也可称为植物性神经系统或内脏神经系统。实际上,自主神经系统还是接受中枢神经系统的控制的,并不是完全独立自主的。按一般惯例,自主神经系统仅指支配内脏器官的传出神经,而不包括传入神经;并将其分成交感神经和副交感神经两部分。(图10-40)
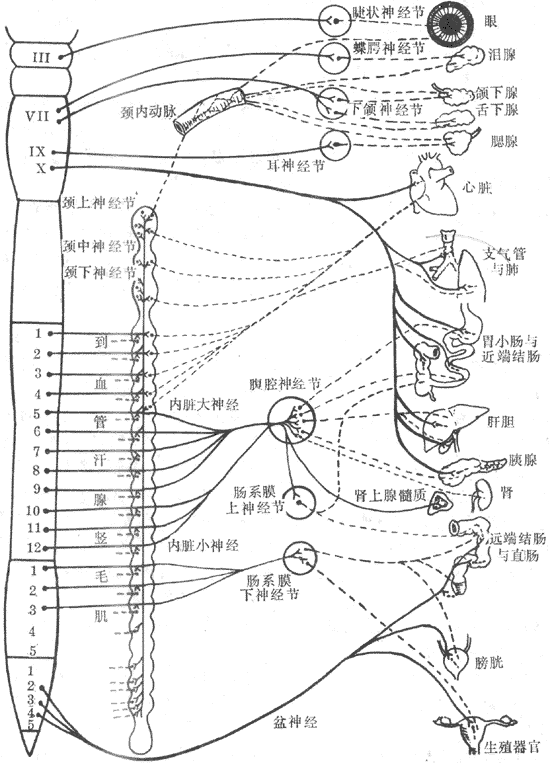
图10-40 自主神经分布示意图
(一)交感和副交感神经的特征
从中枢发出的自主神经在抵达效应器官前必须先进入外周神经节(肾上腺髓质的交感神经支配是一个例外),此纤维终止于节仙神经元上,由节内神经元再发现纤维支配效应器官。由中枢发出的纤维称为节前纤维,由节内神经元发出的纤维称为节后纤维。节前纤维性有髓鞘B类神经纤维,传导速度较快;节后纤维性无髓鞘C类神经纤维,传导速度较慢。交感神经节离效应器官较远,因此节前纤维短而节后纤维长;副交感神经节离效应器官较近,有的神经节就在效应器官壁内,因此节前纤维长而节后纤维短。
交感神经起自脊髓胸腰段的外侧柱。副交感神经的起源比较分解,其一部分起正脑干的缩瞳核、上唾液核、下唾液核、迷走背核、疑核,另一部分起自脊髓骶部相当于侧角的部位。交感神经的全身分布广泛,几乎所有内脏器官都受它支配;而副交感神经的会布较局限,某些器官不具有副交感神经支配。例如,皮肤和肌肉内的血管、一般的汗腺、竖毛肌、肾上腺髓质、肾就只有交感神经支配。
刺激交感神经的节前纤维,反应比较弥散;刺激副交感神经的节前纤维,反应比较局限,因为一根交感节前纤维往往和多个节内神经元发生突触联系,而副交感神经则不同。例如,猫颈上神经节内的交感节前与节后纤维之比为1:11-17,睫状神经节内的副交感节前与节后纤维之比为1:2。
通过荧光组织化学的研究,发现哺乳动物中,交感神经节后纤维交不都是支配效应器官细胞的。在心脏和膀胱中,少量交感神经节后纤维支配器官壁内的神经节细胞;在胃和小肠中,多数交感神经节后纤维支配器官壁内的神经节细胞。由此看来,交感和副交感神经的相互作用,可以发生在器官壁内神经节细胞水平上,而不一定发生在效应器官细胞水平上。
(二)交感和副交感神经系统的功能
自主神经系统的功能在于调节心肌、平滑肌和腺体(消化腺、汗腺、部分内分泌腺)的活动(表10-7)。除少数器官外,一般组织器官都接受交感和副交感的双重支配。在具有双重支配的器官中,交感和副交感神经的作用往往具有拮抗的性质。例如,对于心脏,迷走神经具有抑制作用,而交感神经具有兴奋作用;对于小肠平滑肌,迷走神经具有增强其运动的作用,而交感神经却具有抑制作用。这种拮抗性使神经系统能够从正反两个方面调节内脏的活动,拮抗作用的对立统一是神经系统对内脏活动调节的特点。在一般情况下,交感神经中枢的活动和副交感神经中枢的活动是对立的,也就是说当交感神经系统活动相对加强时,副交感神经系统活动就处于相对减退的地位,而在外周作用方面却表现协调一致。但是,在某些情况下,也可出现交感和副交感神经系统活动都增强或都减退,然而两者间必有一个占优势。在某些外周效应器上,交感和副交感神经的作用是一致的,例如唾液腺的交感神经和副交感神经支配都有促进分泌的作用;但两者的作用也有差别,前者的分泌粘稠,后者的分泌稀薄。
表10-7 自主神经的主要功能
| 器官 | 交感神经 | 副交感神经 |
| 循环器官 | 心跳加快加强 腹腔内脏血管、皮肤血管以及分布于唾液腺与外生殖器官的血管均收缩,脾包囊收缩,肌肉血管可收缩(肾上腺素能)或舒张(胆碱能) | 心跳减慢,心房收缩减弱 部分血管(如软脑膜动脉与分布于外生殖器的血管等)舒张 |
| 呼吸器官 | 支气管平滑肌舒张 | 支气管平滑肌收缩,促进粘膜腺分泌 |
| 消化器官 | 分泌粘稠唾液,抑制胃肠运动,促进括约肌收缩,抑制胆囊活动 | 分泌稀薄唾液,促进胃液、胰液分泌,促进胃肠运动和使括约肌舒张,促进胆囊收缩 |
| 泌尿生殖器官 | 促进肾小管的重吸收,使逼尿肌舒张和括约肌收缩,使有孕子宫收缩,无孕子宫舒张 | 使逼尿肌收缩和括约肌舒张 |
| 眼 | 使虹膜辐射肌收缩,瞳孔扩大使睫状体辐射状肌收缩,睫状体增大 使上眼睑平滑肌收缩 | 使虹膜环形肌收缩,瞳孔缩小 使眼下状体环形肌收缩,睫状体环缩小 促进泪腺分泌 |
| 皮肤 | 竖毛肌收缩,汗腺分泌 | |
| 代谢 | 促进糖原分解,促进肾上腺髓质分泌 | 促进胰岛素分泌 |
自主神经对效应器的支配,一般具有持久的紧张性作用,例如,切断支配心脏的迷走神经,则心率增加,说明心迷走神经本来有紧张性冲动传出,对心脏具有持久的抑制作用;切断心交感神经,则心率关慢,说明心交感神经也有紧张性冲动传出。又如,切断支配虹膜的副交感神经,则瞳孔散大;切断其交感神经,则瞳孔缩小,也说明自主神经的活动具有紧张性。自主神经中枢具有紧张性冲动传出的原因是多方面的,其中有反射性和体液性原因。例如来自主动脉弓和颈动脉窦区域的压力和化学感受器的传入冲动,对维持自主神经的紧张性活动有重要作用;而中枢神经组织内CO2浓度,对维持交感缩血管中枢的紧张性活动与有重要作用。
自主神经的外周性作用与效应器本身的功能状态有关。例如,刺激交感神经可引致动物无孕子宫的运动受到抑制,而对有孕子宫却可加强其运动(因为无孕与有孕子宫的受体不一样);又如,胃幽门如果原来处于收缩状态,则刺激迷走神经使之舒张,如原来处于舒张状态,则刺激迷走神经使之收缩。
交感神经系统的活动一般比较广泛,常以整个系统参与反应。例如,当交感神经系统发生反射性兴奋时,除心血管功能亢进外,还伴有瞳孔散大、支气管扩张、胃肠活动抑制等反应。有人认为,交感神经系统作为一个完整的系统进行活动时,其主要作用在于促使运动机体能适应环境的急聚变化。在剧烈肌肉运动、窒息、失血或冷冻等情况下,机体出现心率加速、皮肤与腹腔内脏血管收缩、血液贮存库排出血液以增加循环轿量、红细胞计数增加、支气管扩张、胆糖原分解加速以及血糖浓度上升、肾上腺素分泌增加等现象,这些现象大多是由于交感神经系统活动亢进所造成的。所以,交感神经系统在环境争聚变化的条件下,可以动员机体许多器官的潜在力量,以适应环境的急变。交感神经系统活动具有广泛性,但并不意味着毫无选择性,实际上交感神经系统发生反射反应时,各部位的交感神经活动还是有判别的。例如,失血后的开始10分钟内,交感神经传出的活动增加,主要表现为心脏活动增强与腹腔内脏血管的收缩,而其他反应就明显;又如,加温刺激下丘脑引致体温调节反应时,皮肤血管的交感神经活动减弱而使皮肤血流增加,但内脏血管的交感神经活动却增强。这些都说明,交感神经系统的反应还是具有相对选择性的。
副交感神经系统的活动,不如交感神经系统的活动那样广泛,而是比较局限的。其整个系统的活动主要在于保护机体、休整恢复、促进消化、积蓄能量以及加强排泄和生殖功能等方面。例如,心脏活动的抑制,瞳孔缩小避免强光的进入,消化道功能增强以促进营养物质吸收和能量补给等,这些都是副交感神经积蓄能量和保护机体的例子。
前文已述及自主神经末梢是通过释放递质而发挥作用的,副交感神经节后纤维和支配汗腺的交感神经节后纤维的递质均为乙酰胆碱,且均属于M样作用;当有机磷中毒时,由于胆碱酯酶失去活性,乙酰胆碱不能被水解而失活,这时就会出现广泛的副交感神经系统兴奋的症状(支气管痉挛、瞳孔缩小、流涎、大小便失禁等),同时大汗淋漓。这些症状均可被大剂量M受体阻断剂(阿托品)所解除,起到抢救的作用。但是,阿托品并不能恢复胆碱酯酶的活性,也不能解除乙酰胆碱N样作用的症状(如骨骼肌颤动),因此抢救时要和胆碱酯酶复活剂(解磷定、氯磷定)联合使用,才能收到更好效果。
二、脊髓对内脏活动的调节
交感神经和部分副交感神经发源于脊髓的外侧柱及相当于外侧柱的部位,因此脊髓可以成为内脏反射活动的初级中枢。在脊髓颈第五节段以上离断的动物,脊休克过去以后,血压可以上升恢复到一定水平,说明脊髓中枢可以完成基本的血管张力反射,以维持血管的紧张性,保持一定的外周阻力;同时还可具有反射性排尿和排粪的能力,说明基本的排尿反射与排便反射可以脊髓中枢内完成。在脊髓高位离断的病人,脊休克过去以后,也可见到血管张力发射、发汗反射、排快活反射、勃起反射的恢复。但是,这种反射调节功能是初级的,不能很好适应生理功能的需要。例如,当由平卧位转成站立时,病人就感到头晕;因为这时体位性血压反射的调节能力很差,脊髓以上的心血管中枢活动已不能控制脊髓的初级中枢,血管的外周阻已不能及时发生改变。又如,损害性刺激虽可以引致发汗反射,但由下丘控制的体温调节性发汗功能去消失。再如,基本的排尿反射可以进行,但排尿不能受意识控制,而且排尿也不完全;但在脊髓离断的病人,搔爬骶部皮肤也可以反射性地引致膀胱收缩而排尿,这使病人能够在一定程度上自己掌握排尿反射。
三、低位脑干对内脏活动的调节
由延髓发出的自主神经传出纤维支配头部的所有腺体、心、支气管、喉头、食管、胃、胰腺、肝和小肠等;同时,脑干网状结构中存在许多与内脏活动功能有关的神经元,其下行纤维支配脊髓,调节着脊髓的自主神经功能。因此,许多基本生命现象(如循环、呼吸等)的反射调节在延髓水平已能初步完成。临床观察和动物实验观察证明,延髓由于受压、穿现等原因而受损时,可迅速造成死亡,以致有人称延髓为基本生命中枢,延髓中的心血管功能、呼吸功能、消化功能等反射调节中枢,已分别在有关章节叙述,不再重复。
此外,中脑是瞳孔对光反射的中枢部位。中脑和下丘脑、边缘前脑对自主神经功能的调节是不可分割的。
四、下丘脑
下丘脑大致可分为四区,即前区、内侧区、外侧区与后区(图10-41)。前区的最前端为视前核,严格说来它属于前脑的范畴,稍后为视上核、视交叉上核、室旁核,再后是下丘脑前核,内侧区又称结节区,紧靠着下丘脑前核,其中有腹内侧核、背内侧核、结节核与灰白结节,还有弓状核与结节乳头核。外侧区有分解的下丘脑外侧核,其间穿插有内侧前脑束。后区主要是下丘脑后核与乳头体核。
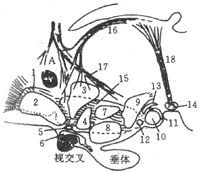
图10-41 下丘脑神经核群示意图
A:前连合 1:外侧视前核 2:内侧视前核 3:室旁核4:下丘脑前核
5:视交叉上核 6:视上核 7:下丘脑背侧核8:下丘脑腹侧内侧核
9:下丘脑后侧核 10,11,12,13:乳头体核群14:中脑之脚间核
15:下丘脑外侧核 16:缰纹 17:穹窿 18:后屈束横贯下丘脑的纤维为内侧前脑束
下丘脑与边缘前脑及脑干网状结构紧密的形态和功能方面的联系,共同调节着内脏的省城。进入下丘脑的传入冲动可来逢边缘前脑、丘脑、脑干网状结构;其传出冲动也可抵达这些部位,还可通过垂体门脉系统和下丘脑-垂体束调节垂体前叶和后叶的活动。垂体门脉是正中隆起(灰白结节的内侧前部)与腺垂体之间的门脉系统;许多含有分泌颗粒的神经末梢终止于正中隆起,其分泌物可通过这一门脉系统到达腺垂体,调节腺垂体的活动,下丘脑-垂体束是由视上核、室旁核和结节发出的神经纤维束,它经垂体柄到达神经垂体,与神经垂体的活动密切相关。
在实验中,曾经观察到电刺激下丘脑的后区可获得血压升高、心率加速、瞳孔散大等交感神经性反应;因此有人认为下丘脑的后部是交感神经中枢,而前部是副交感神经不枢。但这个概念没有得到足够实验事实的支持,已不被公认。现在知道,下丘脑不是单纯的交感和副交感神经中枢,而是较高级的调节内脏活动和其它生理活动联系起来,调节着体温、营养摄取、水平衡、内分泌、情绪反应、生物节律等重要生理过程。
(一)体温调节
哺乳类运动在下丘脑以下部位横切脑干后,即不能保持体温的相对稳定;而在间脑以上切除大脑皮层的动物,体温仍能基本保持相对稳定。可见在间脑水平存在着体温调节中枢。现已肯定,调节体温的中枢在下丘脑。有人认为,体温调节中枢内有些部位能感知温度当血温超过或低于一定水平(这水平称为调定点,正常时约为36.8℃)时,即可通过调节产热和散热活动使体温保持相对稳定。体温调节中枢内的另一些部位对温度变化不敏感,但在温度敏感区的作用下,发出传出冲动以改变与产热和散絷有关器官的活动,从而保持体温的相对稳定。因此下丘脑的体温调节中枢,包括温度感受部分和控制产热和散热功能的整合作用部分。
(二)摄食行为调节
用埋藏电极刺激动物下丘脑外侧区,引致动物多食,而破坏此核后,则动物食欲增大而逐渐肥胖。由此认为,下丘脑外侧区存在摄食中枢(feeding center)。而腹内侧核存在饱中枢(satiety center),后者可以抑制前者活动。用微电极分别记录下丘脑外侧区和腹内侧核的神经元放电,观察到动物在饥锇时,前者放电频率较高而后者放电频率较低;静脉注入葡萄糖后,则前者放频率减少而后者放电频率增多。说明摄食中枢与饱中枢的神经元活动具有相互制约的关系,而且这些神经元对血糖敏感,血糖水平的高低可能调节着摄食中枢和饱中枢的活动。用电渗法(electroosmosis)法将葡萄糖注到腹内侧核神经元旁,也能使神经元放电频率增加,进一步说明饱中枢神经元对葡萄糖敏感。
(三)水平衡调节
水平衡包括水的摄入与排出两个方面,人体通过渴觉引起摄水,而排水则主要取决于肾的活动。损坏下丘脑可引致烦渴与多尿,说明下丘脑对水的摄入与排出调节均关系。
下丘脑控制摄入的区域与上述摄食中枢极为靠近。破坏下丘脑外侧区后,动物除拒食外,饮水也明显减少;刺激下丘脑外侧区某引起部位,则可引致动物饮水增多但是,控制摄水的中枢在确切部位还不清楚,不同动物的实验结果也不一致。
下丘脑控制排水的功能是通过改变抗利尿激素的分泌来完成的。抗利尿激素是由视上核和室旁核的神经元(神经分泌大细胞)合成的,神经分泌颗粒沿下丘脑-垂体束纤维向外周运输(轴浆运输)而贮存于神经垂体。以高渗盐水注入动物的颈内动脉,则能刺激抗利尿激素的分泌;如注入低渗盐水则抑制抗利尿激素的分泌。因此认为,下丘脑内存在着渗透压感受器,它能按血液的渗透压变化来调节抗利尿激素的分泌,此种感受器可能就在视上核和室旁核内。电生理研究观察到,当颈动脉内注入高渗盐水时,视上核内某些神经元放电增多,这一事实支持渗透压感受器就在视上核内的推测。一般认为,下丘脑控制摄水的区域与控制抗利尿激素分泌的核团在功能上是有联系的,两者协同调节着水平衡。
(四)对腺垂体激素分泌的调节
下丘脑内有些神经元(神经分泌小细胞)能正确性调节腺垂体激素分泌的肽类物质,这些物质是:促甲状腺素释放激素、促性腺素释放激素、生长至少释放抑制激素、生长素释放激素、促肾上腺皮质激素释放激素、促黑素细胞激素释放因子、促黑素细胞激素释放抑制因子、催乳素释放因子、催乳素释放抑制因子等。这些肽类物质在全盛后即经轴突运输并分泌到正中隆起,由此经垂体门脉系统到达腺垂体,促进或抑制某种腺垂体激素的分泌。此处,下丘脑还有些神经元对血液中某些激素浓度的变化比较敏感,这咱神经元称为监察细胞;例如,前区的某些神经元对卵巢激素敏感,内侧区的某些神经元对肾上腺皮质激素敏感,另有一些区域的某些神经元对各种垂体促激素很敏感。这些监察细胞在感受血液中激素浓度变化的信息后,可以反馈调节上述肽类物质的分泌,从而更好地控制腺垂体的激素分泌活动。
(五)对情绪生理反应的影响
情绪是人类一种心理现象,但伴随着情绪活动也发生一系列生理变化。这些客观的生理变化,称为情绪生理反应。自主神经系统的情绪反应,可以表现为交感神经系统活动相对亢进的现象,例如猫对痛刺激产生情绪反应时,可以出现心率加速、血压上升、胃肠运动抑制、脚掌出汗、竖毛、瞳孔散大、脾收缩而血液中红细胞计数增加、血糖浓度上升,同时呼吸往往加深加快。人类在发怒情况下,也可见到类似的现象。自主神经系统的情绪反应,在某些情况下也可表现为副交感神经系统活动相对亢进的现象。例如,食物性嗅觉刺激可引致消化液分泌增加和胃肠运动加强,动物发生性兴奋时则生殖器官血管舒张;人类焦急不安可引致排尿排便次数加频,忧虑可引致消化液分泌加多,悲伤则流泪,某些人受惊吓会引致心率减慢。因此,情绪生理反应主要是交感和副交感神经系统活动两者对立统一的改变。持久的情绪活动会造成自主神经系统功能的紊乱。
在间脑水平以上切除大脑的猫,常出现一系列交感神经系统兴奋亢进的现象,并且张牙舞爪,好似正常猫在搏斗时一样,故称之为假怒(sham rage)。平时下丘脑的这种活动受到大脑的抑制而不易表现。切除大脑后则抑制解除,下丘脑的防御反应功能被释放出来,在微弱的刺激下就能激发强烈假怒反应。研究指出,下丘脑内存防御反应区(defense zone),它主要位于下丘脑近中线两旁的腹内侧区。在动物麻醉条件下,电刺激该区可获得骨骼肌的舒血管效应(通过胆碱能交感舒血管纤维),同时伴有血压上升、皮肤及小肠血管收缩、心率加速和其他交感神经性反应。在动物清醒条件下,电刺激该区还可出现防御性行为。此外,电刺激下丘脑外侧区或引致动物出现攻击撕杀行为,电刺激下丘脑背侧区则出现逃避性行为。可见,下丘脑与情绪生理反应的关系很密切。人类下丘脑的疾病也往往伴随着不正常的情绪生理反应。
(六)对生物节律的控制
机体内的各种活动常按一定的时间顺序发生变化,这种变化的节律称为生物节律(biorhygym)。人体许多生理功能都有日周期节律,例如血细胞数、体温、促肾上腺皮质激素分泌等一瑚一个波动周期。身体内各种不同细胞都具有各自的日周期节律,但在自然环境中生活的人体器官组织却表现统一的日周期节律,这说明体内有一个总的控制生物节律的中心,它能使各种位相不同的生物节律统一起来,趋于同步化。研究指出,下丘脑的视交叉上核可能是生物节律的控制中心,在小鼠中观察到视交叉上核神经元的代谢强度和放电活动都表现明确的日周期节律。在胚胎期,当视交叉上核与周围组织还未建立联系时,其代谢和放电活动的日周期节律就已存在。破坏小鼠的视交叉上核,可使原有的日周期节律性活动(如饮水、排尿)的日周期丧失。视交叉上核可通过视网膜-视交叉上核束与视觉感受装置发生联系,因此外环境的昼夜光照变化可影响视交叉上核的活动,从而使体内日周期节律与外环境的昼夜节律同步起来。
五、神经、内分泌和免疫功能的关系
神经与内分泌功能间有密切的关系,近来年的研究发现,神经、内分泌和免疫功能间也有密切的关系,并认为三者共同构成一个完整的调节网络。
(一)神经对免疫功能的作用
神经可以通过两条途径来影响免疫功能,一条是通过神经释放递质来发挥作用,另一条是通过改变内分泌的活动转而影响免疫功能。骨髓、胸腺、淋巴结等免疫器官均有自主神经进入,虽然神经纤维主要是支配血管的,但末梢释放的递质(去甲肾上腺素、乙酰胆碱、肽类)可以通过弥散而作用于免疫细胞。
去甲肾上腺素能抑制免疫反应,免疫细胞上有相应有肾上腺素能受体。乙酰胆碱能增强免疫反应,免疫细胞上的胆碱能受体主要为M型。脑啡肽能增强免疫反应,而β-内啡肽的作用比较多样,有时能促进免疫反应,有时则抑制免疫反应。
神经细胞在特定的条件下也可产生免疫因子,例如在内毒素处理后可产生白细胞介素-1(白介素-1)等。
(二)免疫系统对神经活动的影响
在大鼠实验中观察到,用注入羊红细胞的方法来诱导免疫反应,当抗体生成增多达顶峰时,下丘脑某些神经元的电活动增加1倍以上,提示免疫反应可以改变神经活动。在裸鼠中注入白介素-1,可以使下丘脑有关神经元释放更多的促肾上腺皮持激素释放激素,导致血中促肾上腺皮质激素和糖皮质激素升高几倍,说明白介素-1可以作用于下丘脑神经元。
(三)内分泌系统对免疫功能的影响
促肾上腺皮质激素释放激素能直接促使人外周白细胞(经内毒素预处理后)产生促肾上腺皮质激素和内啡肽。促肾上腺皮质激素具有抑制免疫反应的作用,糖皮质激素一般也具有抑制免疫反应的作用。雌激素、孕激素和雄激素均有抑制免疫功能的作用。促甲状腺素释放激素、促甲状腺素、甲状腺激素均有增强免疫功能的作用。生长激素也有增强免疫功能的作用。
(四)免疫系统对内分泌功能的影响
前文已述及白介素-1能作用于下丘脑而增加促肾上腺皮质激素和糖皮质激素的血中含量。在大鼠中观察到,注入羊红细胞诱导免疫反应达到高峰期间,血中糖皮质激素含量上升而甲状腺激素含量下降,这一机制可能是一种负反馈调节,使免疫反应受到压抑而不致过分。此外,较低浓度的白介素-1能使胰岛B细胞的胰岛素分泌增加。
免疫细胞具有内分泌细胞样功能。免疫细胞分泌折各种免疫因子均为多肽或蛋白质,可以认为免疫因子是免疫细胞产生的内分泌样物质。此外,免疫细胞还可产生一般的激素。例如,巨噬细胞经内毒素外理后能分泌促肾上腺皮质激素、β-内啡肽和脑啡肽,外周淋巴细胞在葡萄糖菌毒素A的刺激下可产生促甲状腺素。
六、大脑皮层对内脏活动的调节
(一)新皮层
在动物实验中电刺激新皮层,除了能引致躯体运动等反应以外,也可引致内脏活动的变化。刺激皮层内侧面4区一定部位,会产生直肠与膀胱运动的变化;刺激皮层外侧面一定部位,会产生呼吸、血管运动的变化;刺激4区底部,会产生消化道运动及唾液分泌的变化。刺激6区一定部位可引致竖毛与出汗,也会引致下肢血管反应的区域也与下肢躯体运动代表区相对应。此外,刺激8区和19区等,除了可引致眼外肌运动外,也可引致瞳也的反应。所有这些结果,说明新皮层与内脏活动有关,而且区域分布和躯体运动代表区的分布有一定的地方。电刺激人类大脑皮层也以觅到类似的结果。
(二)边缘叶
大脑半球内侧面皮层与脑干连接部和胼胝体旁的环周结构,曾被称为边缘叶(图10-42)。这部分结构从进化上是比较古旧的;其最内侧的一环状结构(包括海马、穹窿等)称为古皮层,其较外圈的一环状结构(包括扣带回、海马回等)称为旧皮层。这部分结构曾被认为只与嗅觉联系,而称为嗅脑;但现已明确,其功能远不止这些,而是调节内脏活动的重要中枢。由于边缘叶在结构和功能上和大脑皮层的岛叶、颞极、眶回等,以及皮层下的杏仁核、隔区、下丘脑前核等,是密切相关的,于是有人把边缘叶连同这些结构统称为边缘系统。更有人见到中脑的中央灰质、被盖等也与上述结构有密切的上下行纤维双向联系,于是把中脑的部分结构也包括在边缘系统和概念之中。由此出现了边缘前脑(limbicforebrain)与边缘中脑(limbic midbrain)的概念,前者包括海马、穹窿、海马回、扣带回、杏仁核、隔区、梨状区、岛叶、颞极、眶回等结构,后者指中脑的中央灰质、被盖的中央部分及外侧部、脚间核等。边缘前脑的功能比较复杂,现仅述及其几个方面。
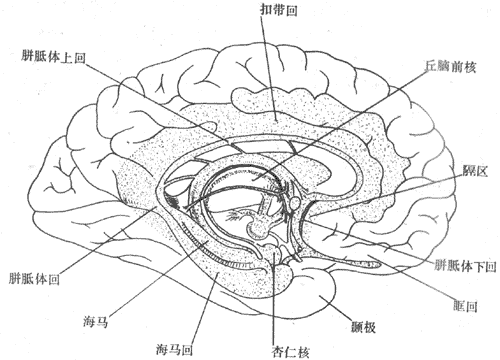
图10-42 大脑内侧面示边缘系统各部位
1.对情绪反应的影响 已知,杏仁核的活动与情绪反应有较密切的关系。杏仁核是由几个核群组成的复合体,从进化来看其中皮层内侧核群比较古老,基底外侧核群进化上较新。皮层内侧核群经终纹与下丘脑腹内侧核有联系,而基底外侧核群经腹侧杏仁传出系统也腹内侧核也有联系;前者有抑制腹内侧核神经元活动的作用,执行者有促进作用,。下丘脑与腹内侧区是防御反应区,因此前者有抑制防御反应作用,后者有易化防御反应的作用。
2.对摄食行为的影响下丘脑腹内侧核区即是防御反应区,也是饱中枢所在的部位。因此杏仁核队了能影响防御反应外,也能影响摄食行为。实验观察到,破坏杏仁核的猫,由于摄食过多而肥胖;用埋藏电极刺激杏仁核的基底外侧核群,可抑制摄食活动;同时记录杏佳核基底外侧核群和下丘脑外侧区(摄食中枢)的神经元放电,可见到两者的自发放电呈相互制约的关系,即当一个放电增我则另一个放电就减少。由此看来,杏仁核的基底外侧核群能易化饱中枢并抑制摄食中枢的活动。此外,刺激隔区,也可观察到相似的结果,即易化饱中枢并抑制摄食中枢的活动。
3.与记忆功能的关系海马与记忆功能有关。由于治疗的需要而手术切除双侧颞中叶的病人,如扣伤了海马及有关结构,则引致近期记忆功能的丧失;手术后对日常遇到的事件丧失记忆力,丧失的程度常决定于损伤部位的大小。临床上还观察到,由于手术切除第三脑室囊肿而损伤了穹窿,也能使患者丧失近期记忆能力;而且还观察到乳头体或乳头体丘脑束的疾患也会导致近期记忆能力的丧失。把这些事实联系起来,可以认为与近期记忆功能有关的神经结构是一个环路结构,即海马→穹窿→下丘脑乳头体→丘脑前核→扣带回→海马,称为海马环路。
4.对其他内脏活动反应的影响刺激边缘前脑不同部位所引起的内脏活动反应是很复杂的,血压可以表现升高或降低,呼吸可以加快或抑制,胃肠运动可以加强或减弱,瞳孔可以扩大或缩小等。例如,刺激扣带回前部可出现患得患失抑制(刺激过强则呼吸加强)、血压下降或上升、心率变慢、胃运动抑制、瞳孔扩大或缩小;刺激杏仁核出现咀嚼、唾液分泌增加、胃酸分泌增加、胃蠕动增加、排便、心率减慢、瞳孔扩大;刺激隔区出现阴茎勃起、血压下降或上升、呼吸暂停或加强。这些运动实验结果,说明边缘前脑的功能和初级中枢的功能不一样;刺激初级中枢扳应比较肯定而一致,而刺激边缘前脑的结果变化较大。可以设想,初级中枢的功能比较局限,活动反应比较单纯;而边缘前脑是许多初级中枢活动的调节者,它能通过促进或抑制各初级中枢的活动,调节更为复杂的生理功能活动,因此活动反应也就复杂而多变。
-
《生理学》 中的相关章节:
……
第十章 神经系统
第一节 神经元活动的一般规律
第二节 反射活动的一般规律
第三节 神经系统的感觉分析功能
第四节 神经系统对躯体运动的调节
第五节 神经系统对内脏活动的调节(当前页)
第六节 脑的高级功能和脑电图
第十一章 内分泌
第一节 概述
第二节 下丘脑的内分泌功能
……