��һ�ڡ�����
�ڷ���ϵͳ�����ڷ����ٺͷֽ������ijЩ��֯�����е��ڷ���ϸ����ɵ�һ��������Ϣ����ϵͳ��������ϵͳ������ϵ�����ϣ���ͬ���ڻ���ĸ��ֹ��ܻ��ά���ڻ�������ȶ���
��������Ҫ���ڷ������д��塢��״�١���״���١������١��ȵ������١��ɹ�������٣�ɢ������֯�����е��ڷ���ϸ���ȽϹ㷺����������ճĤ���ġ������Ρ�Ƥ����̥�̵Ȳ�λ�������ڸ��ָ������ڷ���ϸ�������⣬��������ϵͳ�ڣ��ر���������ڼ����ڷ��ڹ��ܵ���ϸ�������ڷ����ٻ�ɢ���ڷ���ϸ�������ڵĸ�Ч�ܵ�����������ʣ�����֯Һ��ѪҺ���ݶ�������������ã����ֻ�ѧ���ʳ�Ϊ���أ�hormone����
�����ڷ����о��ķ�չ�����ڼ��ش��ݷ�ʽ����ʶ�����롣��������ؾ�ѪҺ������Զ����İ�ϸ�����������ã����ַ�ʽ��ΪԶ����ڣ�telecring����ijЩ���ؿɲ���ѪҺ���䣬������֯Һ��ɢ���������ڽ�ϸ�������ַ�ʽ��Ϊ�Է���(paracrine)������ڷ���ϸ�������ڵļ����ھֲ���ɢ���ַ��������ڸ��ڷ���ϸ�������ӷ������ã����ַ�ʽ��Ϊ�Է���(autocrine)�����⣬����������������ڷ��ڹ��ܵ���ϸ��������ϸ�����ܲ����ʹ����嶯�����ܺϳɺ��ͷż��أ��ʳ����ڷ���ϸ�������Dz����ļ��س�Ϊ����(neurohormone)�����ؿ�����ϸ����ͻ���Ὤ����������ĩ�Ҷ��ͷţ����ַ�ʽ��Ϊ����(neurocrine)��ͼ11-1����
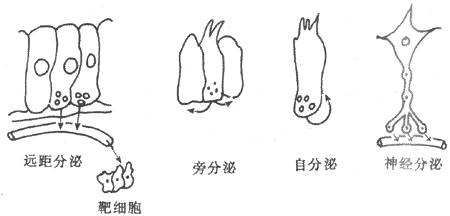
ͼ11-1 ���صĵ��ͷ�ʽ
һ�����صķ���
���ص�����࣬��Դ���ӣ����仯ѧ���ʿɷ�Ϊ�����ࣨ��11-1����
��һ����������
1������͵����ʼ��� ��Ҫ�������Ե����ġ����弤�ء��ٴ��弤�ء��ȵ��ء���״���ټ��ء��������Լ�θ�����صȡ�
2�����༤�� �����������ء�ȥ���������غͼ�״�ټ��ء�
��������̴������壩����
��̴���������������Ƥ�ʺ����ٷ��ڵļ��أ���Ƥ�ʴ���ȩ��ͪ���Ƽ��ء��м����Լ��ۼ��صȡ����⣬���̴�����������1��25-����ά����D3Ҳ����Ϊ���ؿ�����
���⣬ǰ�����ع㷺������������֮֯�У��ɻ�����ϩ��ת�����ɣ���Ҫ����֯�ֲ��ͷţ��ɶԾֲ����ܻ���е��ڣ���˿ɽ�ǰ���ٿ���һ��ֲ����ء�
��11-1 ��Ҫ���ؼ��仯ѧ����
| ��Ҫ��Դ | ���� | Ӣ����д | ��ѧ���� |
| ������ | �ټ�״�ټ����ͷż��� | TRH | ���� |
| �����ټ����ͷż��� | GnRH | ʮ�� | |
| �������ͷ����Ƽ��أ��������أ� | GHRIH | ʮ���� | |
| �������ͷż��� | GHRH | ��ʮ���� | |
| ��������Ƥ�Ʋü����ͷż��� | CRH | ��ʮһ�� | |
| �ٺڣ���ϸ���������ͷ����� | MRF | �� | |
| �ٺڣ���ϸ���������ͷ��������� | MIF | �� | |
| �������ͷ����� | PRF | �� | |
| �������ͷ��������� | PIF | ����ģ����� | |
| ��ѹ�أ��������أ� | VP(ADH) | ���� | |
| �߲��� | OXT | ���� | |
| �ٴ��� | ��������Ƥ�ּ��� | ACTH | ��ʮ���� |
| �ټ�״��Ƥ�ʼ��� | TSH | �ǵ��� | |
| ���ݴ̼��� | FSH | �ǵ��� | |
| ���������أ����ϸ���̼��أ� | LH(ICSH) | �� ���� | |
| �ٺڣ���ϸ�������� | MSH | ʮ���� | |
| ������ | GH | ������ | |
| ������ | PRL | ������ | |
| ��״�� | ��״���أ��ĵ����ԭ���ᣩ | T4 | ���� |
| �������ԭ���� | T3 | ���� | |
| ��״��Cϸ�� | ������ | CT | ��ʮ���� |
| ��״���� | ��״���ټ��� | PTH | ������ |
| �ȵ� | �ȵ��� | ������ | |
| �ȸ�Ѫ���� | ��ʮ���� | ||
| �ȶ��� | ��ʮ���� | ||
| ��Ƥ�ʼ��أ���Ƥ�ʴ��� | ��̴� | ||
| ��Ƥ���أ���ȩ��ͪ�� | ��̴� | ||
| ���� | �������� | E | ���� |
| ȥ���������� | NE | ���� | |
| غ�裺����ϸ�� | غͪ | T | ��̴� |
| ֧��ϸ�� | ������ | �� ���� | |
| �ѳ���̥�� | �ƶ��� | E2 | ��̴� |
| ������ | E3 | ��̴� | |
| ��ͪ | P | ��̴� | |
| ̥�� | ��ëĤ�����ټ��� | CG | �ǵ��� |
| ���������� | θ���� | ʮ���� | |
| ����������-����ø�� | CCK-PZ | ��ʮ���� | |
| ����Һ�� | ��ʮ���� | ||
| �ķ� | �ķ��������� | ANP | ��ʮһ����ʮ���� |
| �ɹ��� | �ʺ��� | ���� | |
| ���� | ���ټ��� | ���� |
�����������õ�һ������
������Ȼ����ܶ࣬���ø��ӣ��������ڶ���֯���ӵ������õĹ����У�����ijЩ��ͬ���ص㡣
��һ�����ص���Ϣ����ʹ��
�ڷ���ϵͳ����ϵͳһ�����ǻ����������Ϣ����ϵͳ�������ߵ���Ϣ������ʽ������ͬ������Ϣ������ά�ϴ���ʱ���Ե��ź�Ϊ��Ϣ��Я���ߣ���ͻ������-ЧӦ����ͷ������ź�Ҫת��Ϊ��ѧ�źţ����ڷ���ϵͳ����Ϣֻ�ǰѻ�ѧ����ʽ��������������ϸ����ϸ��֮�������Ϣ���ݡ����������ּ��أ���ֻ�ܶ�ϸ�����������������ǿ����������ã������书�ܻ�����磬�����شٽ�������������״�ټ�����ǿ��л���̣��ȵ��ؽ���Ѫ�ǵȡ�����Щ�����У����ؼȲ������ӳɷ֣�Ҳ�����ṩ�������������š���ʹ�������ã���������Ϣ���ݸ�����֯��������ǿ�������ϸ����ԭ�е��������������̵����á�
�������������õ����������
�����ͷŽ���ѪҺ�����͵�ȫ��������λ����Ȼ�������������֯��ϸ���й㷺�Ӵ������д˼���ֻ������ijЩ���١���֯��ϸ�������Ϊ�������õ������ԡ�������ѡ�����õ����١���֯��ϸ�����ֱ��Ϊ�����١�����֯�Ͱ�ϸ������Щ����רһ��ѡ��������ijһ�ڷ������壬��Ϊ���صİ��١��������õ����������ϸ���ϴ�������ü��ط��������Խ�ϵ������йء�����͵����ʼ��ص���������ڰ�ϸ��Ĥ�ϣ�����̴��������״�ټ��ص�������λ��ϸ������ϸ�����ڡ������������ʶ���������Խ�ϣ�����ϸ���ڸ��ӵķ�Ӧ���Ӷ�������һ��������ЧӦ����Щ�������õ������Ժ�ǿ��ֻ������ijһ���٣���ټ�״�ټ���ֻ�����ڼ�״�٣���������Ƥ�ʼ���ֻ������������Ƥ�ʣ�����������ټ���ֻ���������ٵȡ���Щ����û���ض��İ��٣������ñȽϹ㷺���������ء���״�ټ��صȣ����Ǽ�����ȫ������֯ϸ���Ĵ�л���̶����ӵ������ã����ǣ���Щ����Ҳ����ϸ������Ӧ�����϶������õġ�
���������صĸ�Ч������Ŵ�����
������ѪҺ�е�Ũ�ȶ��ܵͣ�һ������Ħ����n mol/L��,������ƤĦ����p mol/L������������Ȼ���صĺ������������������ţ���1mg�ļ�״�ټ��ؿ�ʹ�������Ӳ�����Լ4200000 J���������������������Ϻ���ϸ���ڷ���һϵ��ø�ٷŴ����ã�һ����һ�����Ŵ�Ч�����γ�һ��Ч�ܼ���������ŵ�ϵͳ���ݹ��ƣ�һ�����ӵ��ȸ�Ѫ����ʹһ�����ӵ������ỷ��ø�����ͨ��cAMP-����ø���ɼ�����10000�����ӵ����ữø�����⣬һ�����ӵĴټ�״�ټ����ͷż��أ���ʹ�ٴ����ͷ�ʮ������ӵĴټ�״�ټ��ء�0.1��g�Ĵ�������Ƥ�ʼ����ͷż��أ��������ٴ����ͷ�1��g��������Ƥ�ּ��أ�����������������Ƥ�ʷ���40��g��Ƥ�ʼ��أ��Ŵ���400�����ݴ˲�������Ѫ�еļ���Ũ����ͣ���������ȴ�dz����ԣ�������Һ�м���Ũ��ά����Ե��ȶ����Է��Ӽ��ص������������ü�Ϊ��Ҫ��
���ģ����ؼ�������
�����ּ��ع�ͬ����ijһ������ĵ���ʱ�������뼤��֮������������Эͬ���û������ã����ά���书�ܻ������ȶ�������Ҫ���á����磬�����ء��������ء���Ƥ�ʼ��ؼ��ȸ�Ѫ���أ���Ȼʹ�õĻ��ڲ�ͬ�����������Ѫ�ǣ�������ЧӦ����Эͬ���ã��෴���ȵ������Խ���Ѫ�ǣ����������ص�����ЧӦ�������á���״���ټ�����1��12-����ά����D3��Ѫ�Ƶĵ������ศ��ɵģ������������������á�����֮���Эͬ�����������õĻ��ƱȽϸ��ӣ����Է���������ˮƽ��Ҳ���Է������������Ϣ���ݹ��̣�������ϸ����ø�ٷ�Ӧ��ijһ���ڡ����磬��״�ټ��ؿ�ʹ������֯�����ġ��Եȣ���-�����������������ӣ���߶������Ӱ��������ԣ���ǿ��ЧӦ����ͪ��ȩ��ȩ������ˮƽ�����������ã���Ȼ��ͪ��ȩ��ͪ��������Խ�С��������ͪŨ������ʱ�������ȩ��ͪ����ͬһ���壬�Ӷ�����ȩ��ͪ����ˮ�δ�л�����á�ǰ�л��أ�PGI2����ʹѪС����cAMP���࣬�Ӷ�����ѪС��ۼ����෴��Ѫ˨��A2��TXA2��ȴ��ʹѪС����cAMP���٣��ٽ�ѪС��ľۼ���
���⣬�еļ��ر���������ֱ�Ӷ�ijЩ���١���֯��ϸ����������ЧӦ��Ȼ���������ڵ������£���ʹ��һ�ּ��ص�����������ǿ��������һ�ּ����е�����֧�����á����������Ϊ�������ã�permissive action������Ƥ�ʼ��ص����������������Եģ������ļ���Ѫ��ƽ������Ԫ�������ã����ǣ���������Ƥ�ʼ����д��ڣ������Ӱ����ܺܺõط��Ӷ���Ѫ�ܵĵ������á������������õĻ��ƣ�������δ��ȫ�������ȥ��Ϊ������������������Ƥ�ʼ�������������-O-����λø��ʹ�����Ӱ��������ʼ��������������Ӱ�������ǿ������ͨ�������������ˮƽ���о���Ҳ���Ե�������鵼��ϸ����ס�����ݹ��̣���Ӱ�������ỷ��ø�Ļ����Լ�cAMP�����ɵȡ�
�����������õĻ���
������Ϊ��Ϣ�������ϸ���ϵ������Ϻ���ΰ���Ϣ���ݵ�ϸ���ڣ������������Ĵ��۸��ӵķ�Ӧ���̣����ղ���ϸ������ЧӦ�Ļ��ƣ�һֱ���ڷ���ѧ���������о�����Ҫ����һ��ʮ���������ŷ�������ѧ�ķ�չ�����ڼ������û��Ƶ��о��������Ѹ�ٽ�չ�����Ϸḻ�������˹��ڼ������û��Ƶ�����ѧ˵�����ذ��仯ѧ���ʷ�Ϊ�������溬�����غ���̴����أ������༤�������û���Ҳ��ȫ��ͬ���ֱַ�������
��һ���������������û�����ڶ���ʹѧ˵
�ڶ���ʹѧ˵��Sutherland����1965��������ġ�Sutherlandѧ�����о���ԭ�ͽ��һ����������ø�����ữø�Ļ���ʱ�����ȸ�Ѫ�������������ؿ�ʹ���Ƚ���APT��Mg2+�������ỷ��ø��adenylate cyclase,AC���������²���һ�������ʣ��������ʾ��м���������ø�Ӷ�����ԭ�ͽ�����á�ʵ��֤�������ǻ�-�����գ�cyclic AMP,cAMP������Mg2+���ڵ������£������ữø�ٽ�ATPAת��ΪcAMP��CAMP���������ø��phosphodiesterase���������£�����Ϊ5�@AMP�����һ������cAMP֮�����ܼ������ữø��������cAMP��������һ��ø��������cAMP�ĵ���ø��cAMP-dependent protein kinase,cAMP-PK,PKA������ɵġ�
Sutherland�ۺ���Щ��������ڶ���ʹѧ˵������Ҫ���ݰ������ټ����ǵ�һ��ʹ���������ϸ��Ĥ�Ͼ������幹�͵�רһ�������ϣ��ڼ����������Ϻ���Į�ϵ������ỷ��øϵͳ������mg2+Ш���ڵ������£������ỷ��ø��ʹATPת��ΪcAMP��cAMP�ǵڶ���ʹ����Ϣ�ɵ�һ��ʹ���ݸ��ڶ���ʹ����cAMP��ʹ���Եĵ���ø��PKA�����PKA���������ǵ�λ���������ǵ�λ����ǵ�λ��CAMP��PKA�ĵ����ǵ�λ��ϣ����µ����ǵ�λ����������ʹPKA�����ϸ���ڶ��ֵ����ʷ������ữ��Ӧ������һЩ���������ữ���Ӷ������ϸ����������������Ӧ��ͼ11-2����
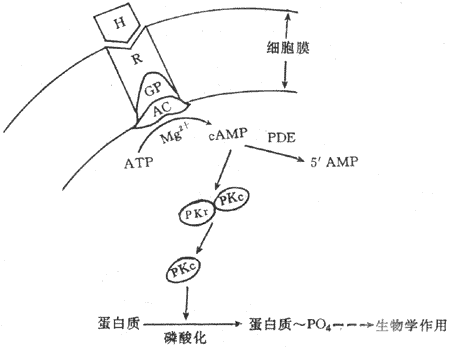
ͼ11-2 �����������û���ʾ��ͼ
H������ R������ GP��G���� AC�������ỷ��øPDE������ ����ø
RKr :����ø�����ǵ�λ PKc:����ø���ǵ�λ
��cAMPΪ�ڶ���ʹѧ˵��ָ�����ƶ��˼������û��Ƶ��о�����Ѹ�����뷢չ�����������о����ϱ�����cAMP������Ψһ�ĵڶ���ʹ��������Ϊ�ڶ���ʹ�Ļ�ѧ���ʻ���cGMP�������ἡ�����������͡�Ca2+�ȡ����⣬����ϸ������������ڡ������ỷ��ø����ơ�����øC�����õȷ�����о���ȡ���˺ܴ��չ���ָ������£�
1������������������ ������Ĥ�����Ϊ�ǵ��ף���ṹһ���Ϊ�����֣�ϸ��Ĥ�����Ρ���Ĥ���ֺ�ϸ��Ĥ�����Ρ�ϸ��Ĥ�����κ��������ǻ�����ʶ���ز���֮��ϵIJ�λ�����ط��ӺͰ�ϸ������ı��֣���������ԳƵĹ��ܻ��Ź��ɼ�Ϊ���Ӷ��ֿɱ�����幹�͡����غ����������յ����ı䱾���Ĺ�������Ӧ�Է��Ĺ��ͣ����Ϊ���������巢��רһ�Խ���ṩ�����ʻ�����
����������Ľ������Ϊ������affinity����һ����˵�����������Ǽ������õĵ�һ�������������뼤�ص�����ѧ��������һ�£������ص���������������϶������ּ��ص����ã��෴ȴ��ϼ������������ϡ�ʵ��֤�����������������������ı仯�������ı䣬�綯�������ڵIJ�ͬ�Σ��ѳ�����ϸ���ϵĵ����ݴ̼��أ�FSH������������Dz���ͬ�ġ�ijһ������������ʱ�����ڽ����������Ҳ�ɳ������͵�����
��������������ı��⣬������Ҳ�ɷ����仯���������ܰ�ϸ��Ĥ���ȵ���������й۲췢�֣��糤��ʹ�ô�������ȵ��أ��������ȵ��������������٣�����Ҳ���ͣ������ȵ��ص������ͺ�����������������ɻָ������������ּ��أ���ټ�״�ټ��ء���ëĤ�����ټ��ء����������ء����ݴ̼��صȣ��������������������ּ���ʹ�������������������ٵ�����Ϊ��˥���ڻ����µ���down regulation0�����µ������Ļ��ƿ����뼤��-���帴������������йء��෴����Щ���أ����ڼ�����Сʱ��Ҳ��ʹ�������������������࣬��Ϊ�������ڻ����ϵ���up regulation����������ء����ݴ̼��ء�Ѫ�ܽ����صȶ����Գ����ϵ������µ����ϵ�����˵����������ϵ�����˵��������ĺϳ��뽵��ڶ�̬ƽ��֮�У�����������һƽ��Ľ�������Ķ����뼤����������Ӧ���Ե��ڰ�ϸ���Լ������������뷴Ӧǿ�ȡ�
2��G��������Ϣ�����е����ü��������������ỷ��ø��ϸ��Ĥ������ֿ��ĵ����ʡ����������ϵIJ�����ϸ��Ĥ������棬�����ջ���ø��Ĥ�İ����棬������֮�����һ�����������õĵ��ڵ��ס����������ϵ��ף�guanine nucleotide-bindingregulatory protein�������G���ס�G�����ɦ����ºͦ������ǵ�λ��ɣ����ǵ�λ������������λ�㡣��G�����Ͻ�ϵ�������ΪGTPʱ����������ã�����G�����ϵ�GTPˮ��ΪGDPAʱ��ʧȥ���ԡ���������������ʱ������������G���Ħ��ǵ�λ��ϣ�����ʹ����¡����ǵ�λ���룬���ܶ������ỷ��ø����������á�
G���ɷ�Ϊ�˷���G���ף�Gs����������G���� Gi��Gs�������Ǽ��������ỷ��ø���Ӷ�ʹcAMP�������ࣻGi�������������������ỷ��ø�Ļ��ԣ�ʹcAMP���ɼ��١����������ϸ��Ĥ�ļ�������Ҳ�ɷ�Ϊ�˷��ͣ�Rs���������ͣ�Ri�����֣����Ƿֱ����˷��Լ��أ�Hs���������Լ���(Hi)������ϣ����ֱ�����Gs��Gi����ͨ����������������ỷ��øʹcAMP���ӻ���ٶ��������á�
3�������ἡ���Ͷ�������Ϊ�ڶ���ʹ����Ϣ����ϵͳ���ຬ����������cAMPΪ�ڶ���ʹ����ϸ�����ܻ�ģ�����Щ�������ص�������Ϣ������cAMPΪý����д��ݣ����ȵ��ء��߲��ء������ء�ijЩ�����Ե����ĺ��������ӵȡ�ʵ��֤������Щ����������Ĥ�������������ϸ��Ĥ��֬������ת���Ϊ�����ἡ��(inositol-1,4,5,triphosphate,IP3)�Ͷ�������(diacylglycerol,DG)�������°�����Ca2+Ũ�����ߡ����������������IP3��DG�����ǵڶ���ʹ��ѧ˵�������ǵ����ӣ����ҵõ�Խ��Խ�ҵ�ʵ��֤ʵ����һѧ˵��Ϊ���ڼ��ص������£�����ͨ��G���Ľ鵼������ϸ��Ĥ�ڵ���֬øC(phosphinositol-specificphospholipase C.PLC)����ʹ����֬��������PI���������ữ���ɵ���֬����������PIP2���ֽ⣬����IP3��DG��DG���ɺ�������Ĥ�У�IP3������������δ�ܵ���������ʱ��ϸ��Ĥ���������������DG��ϸ����IP3�ĺ���Ҳ����ֻ����ϸ��3�ܵ���Ӧ��������ʱ���ż���PIP2�Ľ��⣬��������IP3��DG��IP3�������Ǵ�ʹϸ����Ca2+������ͷ�Ca2+���������ϸ����Ca2+��Ҫ���������������������С�ʵ��֤����IP3����Ca2+���ͷ������������������������壬��Ϊ��������Ĥ����IP3���壬IP3���������������Ϻ���Ca2+ͨ����ʹCa2+���������н��������IP3�շ�Ca2+��Ա�������Ӧ�������ݶ̵��������ͷ�Ca2+���������Ca2+�ͷ��շ����ýϳ���ϸ����Ca2+���������°�����Ca2+Ũ�����ӡ�Ca2+��ϸ���ڵĸƵ����ף�calmodulin,CaM����Ϻɼ����ø���ٽ����������ữ���Ӷ�����ϸ���Ĺ��ܻ��
DG��������Ҫ�����������Լ����øC��protein kinase C,PKC��PKC�ļ���������Ca2+�Ĵ��ڡ������PKC��PKAһ����ʹ���ֵ����ʻ�ø�������ữ��Ӧ����������ϸ��������ЧӦ�����⣬DG�Ľ�����ﻨ����ϩ���Ǻϳ�ǰ�����ص�ԭ�ϣ�������ϩ����ǰ�����صĹ��������ֲ��������ỷ��ø�ļ���ٽ�cGMP�����ɡ�CGMP��Ϊ��һ�ֿ��ܵĵڶ���ʹ��ͨ�������øG(PKG)���ı�ϸ���Ĺ��ܣ�ͼ11-3����
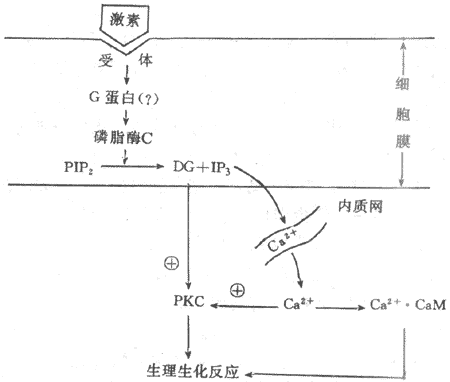
ͼ11-3 ��֬��������Ϣ����ϵͳʾ��ͼ
PIP2����֬�������� DG���������� IP3�������ἡ��PKC������øC CaM:�Ƶ�����
��������̴��������û�����������ѧ˵
��̴����صķ���С����������Ϊ300���ң�����֬���ԣ���˿���ϸ��Ĥ����ϸ�����ڽ���ϸ��֮������������Ӱ����������������ã��ʰѴ������û��Ƴ�Ϊ��������ԭ�������Ϊ�������ѧ˵��
��һ���Ǽ�������������ϣ��γɼ���-�������帴����ڰ�ϸ�����д�������̴��������壬�����ǵ����ʣ�����Ӧ���ؽ���ص���רһ��ǿ�����Դ����磬�ӹ���֯�����Ĵƶ�����������17��-�ƶ�����ϣ���������17��-�ƶ�����ϡ���������������Դ�С�뼤�ص�����ǿ����ƽ�еġ����Ұ�������ĺ���Ҳ������ٵĹ���״̬�ı仯�������ı䡣�����ؽ���ϸ��������������Ϻ����嵰�������ͱ仯���Ӷ���������-�������帴�����ý�����ڵ��������ɰ���ת�������ڡ��ڶ�����������������ϣ��γɼ���-�����帴����Ӷ�����DNA��ת¼���̣������µ�mRNA���յ������ʺϳɣ�������Ӧ������ЧӦ��ͼ11-4����
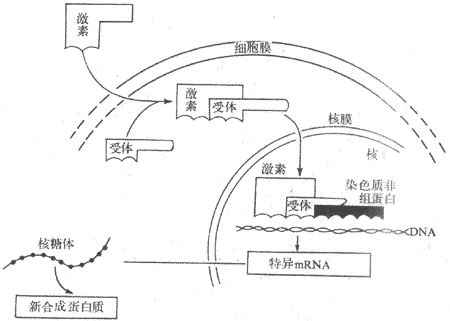
ͼ11-4 ��̴��������û���ʾ��ͼ
���������ڻ��̼����ķ�չ��Ӧ�ã�������̴����صĺ�������Ľṹ�Ѿ����������������ض�ת¼��������õĵ��ף����������̴����صĿ��ơ���������Ҫ���������ܽṹ���ؽ�Ͻṹ��DNAA�ṹ�ṹ���ת¼��ǿ�ṹ��һ�������������ϣ�����ķ��ӹ������ı䣬��¶�������ڷ����ڲ���DNA��Ͻṹ��ת¼��ǿ�ṹ��ʹ����DNA��ϣ��Ӷ�������ǿת¼��ЧӦ�����⣬���μ�ʵ�����ϱ�������DNA��Ͻṹ�������һ���������еİ�����Ƭ�ϣ������Ž鵼�������帴������Ⱦɫ�����ض��IJ�λ���ϣ����Ӻ˶�λ�źŵ����á�
��״�ټ��������������أ��������û���ȴ����̴��������ƣ����ɽ���ϸ���ڣ�������������������ϼ�������ڣ���������ϵ��ڻ�����
Ӧ��ָ�����������ؿ�������ת¼�뷭��ζ�Ӱ�쵰���ʵĺϳɣ�����������̴�����Ҳ����������ϸ��Ĥ����������ѧ˵���Խ��͵�ij��������
-
������ѧ�� �е�����½ڣ�
������
���������ڡ���ϵͳ�ĸо���������
�������Ľڡ���ϵͳ�������˶��ĵ���
��������ڡ���ϵͳ�������ĵ���
���������ڡ��Եĸ����ܺ��Ե�ͼ
��ʮһ�¡��ڷ���
������һ�ڡ���������ǰҳ��
�����ڶ��ڡ������Ե��ڷ��ڹ���
���������ڡ�����
�������Ľڡ���״��
��������ڡ���״���ٺͼ�״��Cϸ��
������